 |


Гладкая мышечная ткань нейрального происхождения
|
|
|
|
Миоциты этой ткани развиваются из клеток нейрального зачатка в составе внутренней стенки глазного бокала. Тела этих клеток располагаются в эпителии задней поверхности радужки. Каждая из них имеет отросток, который направляется в толщу радужки и ложится параллельно ее поверхности. В отростке находится сократительный аппарат, организованный так же, как и во всех гладких миоцитах. В зависимости от направления отростков (перпендикулярно или параллельно краю зрачка) миоциты образуют две мышцы — суживающую и расширяющую зрачок.
39. Исчерченная мышечная ткань: источник развития, строение, иннервация. Сокращение мышечного волокна, типы мыш волокон. Регенерация.
Поперечнополосатые мышечные ткани
Имеется две основные разновидности поперечнополосатых (исчерченных) тканей — скелетная мышечная ткань и сердечная мышечная ткань.
Скелетная мышечная ткань
Гистогенез
Источником развития элементов скелетной (соматической) поперечнополосатой мышечной ткани являются клетки миотомов — миобласты. Одни из них дифференцируются на месте и участвуют в образовании так называемых аутохтонных мышц. Другие клетки мигрируют из миотомов в мезенхиму. Они уже детерминированы, хотя внешне не отличаются от других клеток мезенхимы. Их дифференцировка продолжается в местах закладки других мышц тела.
В ходе дифференцировки возникают две клеточные линии. Клетки одной из линий сливаются, образуя удлиненные симпласты — мышечные трубочки (миотубы). В них происходит дифференцировка специальных органелл — миофибрилл. В это время в миотубах отмечается хорошо развитая гранулярная эндоплазматическая сеть. Миофибриллы сначала располагаются под плазмолеммой, а затем заполняют большую часть миотубы. Ядра, напротив, из центральных отделов смещаются к периферии. Клеточные центры и микротрубочки при этом полностью исчезают. Гранулярная эндоплазматическая сеть редуцируется в значительной степени. Такие дефинитивные структуры называют миосимпластами.
|
|
|
Клетки другой линии остаются самостоятельными и дифференцируются в миосателлитоциты (или миосателлиты). Эти клетки располагаются на поверхности миосимпластов.
Строение
Основной структурной единицей скелетной мышечной ткани является мышечное волокно, состоящее из миосимпласта и миосателлитоцитов, покрытых общей базальной мембраной.
Длина всего волокна может измеряться сантиметрами при толщине всего 50—100 мкм. Комплекс, состоящий из плазмолеммы миосимпласта и базальной мембраны, называют сарколеммой.
Миосимпласт имеет множество продолговатых ядер, расположенных непосредственно под сарколеммой. Их количество в одном симпласте может достигать нескольких десятков тысяч. У полюсов ядер располагаются органеллы общего значения — аппарат Гольджи и небольшие фрагменты гранулярной эндоплазматической сети. Миофибриллы заполняют основную часть миосимпласта и расположены продольно.
Саркомер — это структурная единица миофибриллы. Каждая миофибрилла имеет поперечные темные и светлые диски, имеющие неодинаковое лучепреломление (анизотропные A-диски и изотропные I-диски). Каждая миофибрилла окружена продольно расположенными и анастомозирующими между собой петлями агранулярной эндоплазматической сети — саркоплазматической сети, или саркоплазматического ретикулума. Соседние саркомеры имеют общую пограничную структуру — Z-линию (или телофрагму). Она построена в виде сети из белковых фибриллярных молекул, среди которых существенную роль играет альфа-актинин. С этой сетью связаны концы тонких, актиновых, филаментов. От соседних Z-линий актиновые филаменты направляются к центру саркомера, но не доходят до его середины. Филаменты актина объединены с Z-линией и нитями миозина фибриллярными нерастяжимыми молекулами небулина. Посередине темного диска саркомера располагается сеть, построенная из миомезина. Она образует в сечении М-линию, или мезофрагму. В узлах этой М-линии закреплены концы толстых, миозиновых филаментов. Другие их концы направляются в сторону Z-линий и располагаются между филаментами актина, но до самих Z-линий тоже не доходят. Вместе с тем эти концы фиксированы по отношению к Z-линиям растяжимыми гигантскими белковыми молекулами титина.
|
|
|
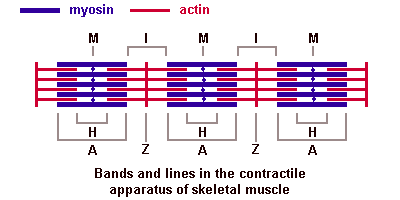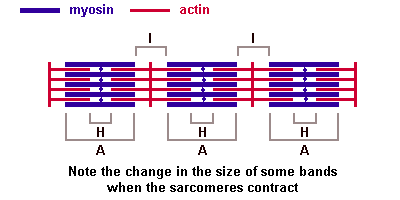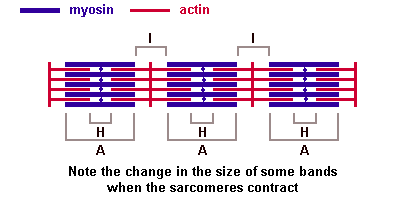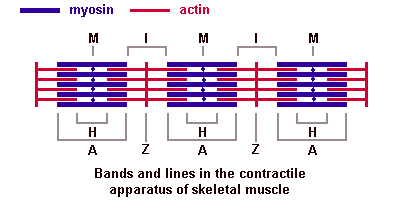 Молекулы миозина имеют длинный хвост и на его конце две головки. При повышении концентрации ионов кальция в области присоединения головок (в своеобразном шарнирном участке) молекула миозина изменяет свою конфигурацию. При этом (поскольку между миозиновыми филаментами расположены актиновые) головки миозина связываются с актином (при участии вспомогательных белков — тропомиозина и тропонина). Затем головка миозина наклоняется и тянет за собой актиновую молекулу в сторону М-линии. Z-линии сближаются, саркомер укорачивается.
Молекулы миозина имеют длинный хвост и на его конце две головки. При повышении концентрации ионов кальция в области присоединения головок (в своеобразном шарнирном участке) молекула миозина изменяет свою конфигурацию. При этом (поскольку между миозиновыми филаментами расположены актиновые) головки миозина связываются с актином (при участии вспомогательных белков — тропомиозина и тропонина). Затем головка миозина наклоняется и тянет за собой актиновую молекулу в сторону М-линии. Z-линии сближаются, саркомер укорачивается.
Альфа-актининовые сети Z-линий соседних миофибрилл связаны друг с другом промежуточными филаментами. Они подходят к внутренней поверхности плазмолеммы и закрепляются в кортикальном слое цитоплазмы, так что саркомеры всех миофибрилл располагаются на одном уровне. Это и создает при наблюдении в микроскоп впечатление поперечной исчерченности всего волокна.
Источником ионов кальция служат цистерны агранулярной эндоплазматической сети. Они вытянуты вдоль миофибрилл около каждого саркомера и образуют саркоплазматическую сеть. Именно в ней аккумулируются ионы кальция, когда миосимпласт находится в расслабленном состоянии. На уровне Z-линий (у амфибии) или на границе А- и I-дисков (у млекопитающих) канальцы сети меняют направление и располагаются поперечно, образуя расширенные терминальные или (латеральные) L-цистерны.
С поверхности миосимпласта плазмолемма образует длинные трубочки, идущие поперечно в глубину клетки (Т-трубочки) на уровне границ между темными и светлыми дисками. Когда клетка получает сигнал о начале сокращения, этот сигнал перемещается по плазмолемме в виде потенциала действия и распространяется отсюда на мембрану Т-трубочек. Поскольку эта мембрана сближена с мембранами саркоплазматической сети, состояние последних меняется, кальций освобождается из цистерн сети и взаимодействует с актино-миозиновыми комплексами (они сокращаются). Когда потенциал действия исчезает, кальций снова аккумулируется в цистернах саркоплазматического ретикулума и сокращение миофибрилл прекращается. Для развития усилия сокращения нужна энергия. Она освобождается за счет АТФ- АДФ-превращений. Роль АТФазы выполняет миозин. Источником АТФ служат главным образом митохондрии, поэтому они и располагаются непосредственно между миофибриллами.
|
|
|
Большую роль в деятельности миосимпластов играют включения миоглобина и гликогена. Гликоген служит источником энергии, необходимой не только для совершения мышечной работы, но и поддержания теплового баланса всего организма. Миоглобин связывает кислород, когда мышца расслаблена и через мелкие кровеносные сосуды свободно протекает кровь. Во время сокращения мышцы сосуды сдавливаются, а запасенный кислород освобождается из миоглобина и участвует в биохимических реакциях.
Миосателлитоциты - это малодифференцированные клетки, являющиеся источником регенерации мышечной ткани. Они прилежат к поверхности миосимпласта, так что их плазмолеммы соприкасаются. Миосателлитоциты одноядерны, их ядра овальной формы и мельче, чем в симпластах. Они обладают всеми органеллами общего значения (в том числе и клеточным центром).
Типы мышечных волокон. Разные мышцы (как органы) функционируют в неодинаковых биомеханических условиях. Поэтому и мышечные волокна в составе разных мышц обладают разной силой, скоростью и длительностью сокращения, а также утомляемостью. Ферменты в них обладают разной активностью и представлены в различных изомерных формах. Заметно различие в них содержания дыхательных ферментов — гликолитических и окислительных.
|
|
|
По соотношению миофибрилл, митохондрий и миоглобина различают белые, красные и промежуточные волокна. По функциональным особенностям мышечные волокна подразделяют на быстрые, медленные и промежуточные. Наиболее заметно мышечные волокна различаются особенностями молекулярной организации миозина. Среди различных его изоформ существуют две основных — «быстрая» и «медленная». При постановке гистохимических реакций их различают по АТФазной активности. С этими свойствами коррелирует и активность дыхательных ферментов. Обычно в быстрых волокнах преобладают гликолитические процессы, они более богаты гликогеном, в них меньше миоглобина, поэтому их называют также белыми. В медленных волокнах, напротив, выше активность окислительных ферментов, они богаче миоглобином, выглядят более красными.
Свойства мышечных волокон меняются при изменении нагрузок — спортивных, профессиональных, а также в экстремальных условиях (таких как невесомость). При возврате к обычной деятельности такие изменения обратимы. При некоторых заболеваниях (мышечные атрофии, дистрофии, последствия денервации) мышечные волокна с разными исходными свойствами изменяются неодинаково. Это позволяет уточнять диагноз, для чего исследуют биоптаты скелетных мышц.
|
|
|
