 |


Основные процессы клеточного метаболизма
|
|
|
|
Содержание
Введение. 3
Понятие и функции клеток. 4
Основные процессы клеточного метаболизма. 11
Заключение. 20
Список использованных источников. 21
Введение
Все живые существа состоят из клеток – маленьких, окруженных мембраной полостей, заполненных концентрированным водным раствором химических веществ. Клетка – элементарная единица строения и жизнедеятельности всех живых организмов (кроме вирусов, о которых нередко говорят как о неклеточных формах жизни), обладающая собственным обменом веществ, способная к самостоятельному существованию, самовоспроизведению и развитию. Все живые организмы либо, как многоклеточные животные, растения и грибы, состоят из множества клеток, либо, как многие простейшие и бактерии, являются одноклеточными организмами. Раздел биологии, занимающийся изучением строения и жизнедеятельности клеток, получил название цитологии. Считается, что все организмы и все составляющие их клетки произошли эволюционным путем от общей преДНКовой клетки.
В нормально функционирующей живой клетке одномоментно протекает множество катализируемых ферментами химических реакций, приводящих к образованию огромного количества разнообразных соединений. В норме обмен веществ в клетке (метаболизм) осуществляется по принципам строжайшей экономии энергии и вещества, что обеспечивается сложнейшей системой регуляции обмена веществ.
Метаболизм представляет собой химические реакции, возникающие с момента поступления в организм питательных веществ до момента выделения во внешнюю среду конечных продуктов этих реакций. Это сложный процесс преобразования потребляемой пищи в жизненную энергию. В метаболизм вовлечены все реакции, протекающие в живых клетках, результатом которых является строительство структур тканей и клеток, т.е. метаболизм можно рассматривать, как процесс обмена в организме веществ и энергии.
|
|
|
Понятие и функции клеток
Понятие о клетках возникло в XVII в., когда впервые стали применять с целью научных наблюдений оптические стекла (линзы). Голландец А. Левенгук, который занимался шлифованием стекол, из любви к этому тонкому искусству впервые начал рассматривать мельчайшие живые существа в капле воды. Он создал первый микроскоп и положил основание научной микроскопии. К новой области увлекательных исследований обратились многочисленные ученые. В результате их работ к 30-м годам XIX столетия накопился обширный материал по изучению микроскопического строения живой природы, на основе которого были сделаны чрезвычайно важные выводы. Установлено было, с одной стороны, что существуют простейшие организмы, представленные всего лишь одной клеткой, с другой стороны, выяснилось, что растения и животные, имеющие сложное строение тела, состоят из огромного числа клеток. При этом оказалось, что все клетки, какому бы организму они ни принадлежали, имеют одинаковые законы строения. Отсюда был сделан важнейший вывод, что все живые существа нашей планеты — и растения, и животные — имеют общее происхождение, общие источники развития в процессе эволюции.

Рисунок 1. а – животная клетка; б – растительная клетка.
1 – оболочка; 2 – протоплазма; 3 – ядро
Строение клетки можно схематически представить следующим образом (рисунок 1). Ее основные части составляют протоплазма, ядро и оболочка. Главной частью клетки является протоплазма. По внешнему виду она представляет вязкую, бесцветную прозрачную жидкость. Химический состав ее очень сложен: сюда входят органические вещества — белки, жиры, углеводы, витамины — и неорганические вещества — вода, минеральные соли. По объему вода составляет 95—98% (!) всей массы клетки; вода с растворенными в ней солями составляет среду, в которой протекают все процессы жизнедеятельности клетки. Из органических веществ важнейшую роль играют белки. Именно белковые молекулы обеспечивают в силу особенности своего строения и химических свойств основной жизненный процесс — обмен веществ. Жиры и углеводы служат главным образом энергетическим материалом: за счет их окисления («сгорания») образуется энергия, необходимая для жизни.
|
|
|
Клетка – это ограниченная активной мембраной, упорядоченная структурированная система биополимеров (белков, нуклеиновых кислот) и их макромолекулярных комплексов, участвующих в единой совокупности метаболических и энергетических процессов, осуществляющих поддержание и воспроизведение всей системы в целом.
В клетках постоянно осуществляется обмен веществ, или метаболизм(от греч. metabole - перемена, превращение), который представляет собой совокупность процессов ассимиляции (реакций биосинтеза сложных биологических молекул из более простых) и диссимиляции (реакций расщепления). В результате диссимиляции освобождается энергия, заключенная в химических связях пищевых веществ. Эта энергия используется клеткой для осуществления различной работы, в том числе и ассимиляции.
Все биохимические реакции в клетке строго структурированы и осуществляются при участии высокоспецифических биокатализаторов - ферментов, или энзимов (от греч. en - в, zyme - брожение, закваска). Ферменты ускоряют реакцию на 10 порядков (в 1010 раз). По общему типу воздействия ферменты подразделяются на 6 классов.
Оксиредуктазы катализируют окислительно-восстановительные реакции, трансферазы участвуют в переносе функциональных групп, гидролазыобеспечивают реакции гидролиза, лиазы - присоединение групп по двойным связям, изомеразы осуществляют перевод соединений в другую изомерную форму, а лигазы связывают молекулярные группировки в цепи.
Все процессы в клетках происходят при участии поступающих в клетку веществ (эндоцитоз) и выведения веществ из клетки (экзоцитоз). Эти процессы, а также внутриклеточный перенос веществ в мембранных пузырьках связаны со слипанием и объединением бислоев мембран (рис. 1). При эндоцитозе слипаются два наружных монослоя плазматической мембраны, при экзоцитозе - два внутренних, обращенных к цитоплазме слоя.
|
|
|
Эндоцитоз. Существует несколько способов эндоцитоза (от греч. endon - внутри, kytos - клетка). При эндоцитозе этот транспорт осуществляется с участием цитолеммы. Кроме того, существуют более сложные способы - пиноцитоз (от греч.pino - пью) и фагоцитоз (от греч. phagos - пожирающий). Обычно под пиноцитозом понимают захват клеткой жидких коллоидных частиц, под фагоцитозом - захват корпускул (более плотных и крупных частиц, а также других клеток). Механизм пино- и фагоцитоза различен.
Пиноцитоз. Для того, чтобы внешние молекулы поступили в клетку, они должны быть сначала связаны рецепторами гликокаликса. Цитолемма вместе с присоединенными извне молекулами впячивается, затем ее края сближаются и смыкаются. В результате от цитолеммы отщепляется пузырек, несущий в себе захваченные молекулы. Такие пузырьки получили название окаймленных. Окаймленные пузырьки беспрепятственно транспортируются в клетке именно к тем участкам цитоплазмы, где должно использоваться их содержимое. Если вещества переносятся через клетку из одной среды в другую без изменений, то этот процесс называют трансцитозом. Путем трансцитоза могут переноситься и белковые молекулы, в частности иммуноглобулины.
Фагоцитоз. Крупные частицы также могут распознаваться рецепторами клетки и захватываться клетками. Выросты клетки охватывают частицу и объединяются над ней. Наружные листки выростов сливаются, замыкая поверхность клетки. Глубокие листки выростов образуют мембрану вокруг поглощенной частицы - формируется фагосома.
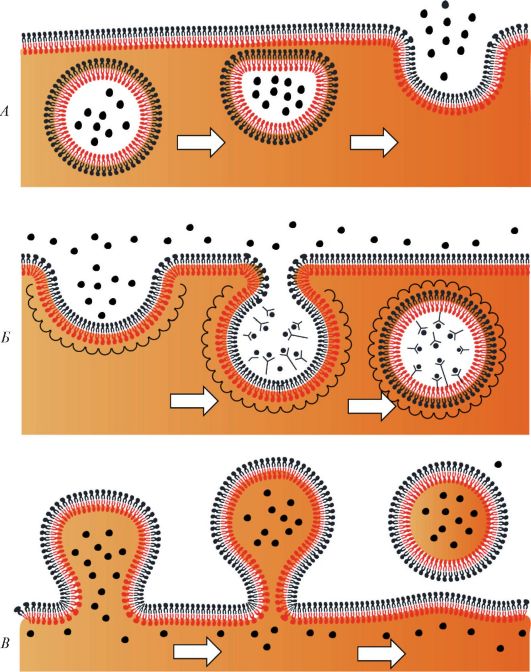
Рисунок 2. Слипание и объединение бислоев (мембран) в трех процессах: в экзоцитозе (А), в эндоцитозе (Б) и в отпочковывании (В). Стрелками показана последовательность этих процессов сливается с лизосомами, в результате чего возникает их комплекс - гетеролизосома (гетеросома).
|
|
|
Способностью к фагоцитозу потенциально обладают все клетки, но в организме лишь некоторые специализируются в этом направлении. Таковы нейтрофильные лейкоциты и макрофаги.
Экзоцитоз. Выведение веществ из клетки осуществляется благодаря нескольким механизмам. Один из них - пассивный транспорт вследствие разности концентраций веществ внутри и вне цитолеммы, другой - активный транспорт. Таким путем выводятся из клетки ионы и мелкие молекулы. Третий механизм обеспечивает выведение крупномолекулярных соединений из клетки.
Крупномолекулярные соединения в виде транспортных пузырьков с участием микротрубочек направляются к клеточной поверхности. Мембрана пузырька сливается с цитолеммой, и содержимое пузырька оказывается за пределами клетки.
Слияние пузырька с цитолеммой может совершаться без каких-либо дополнительных сигналов. Такой экзоцитоз называют конститутивным. Так выводится из клетки большинство продуктов ее собственного метаболизма. Ряд клеток предназначен для синтеза специальных соединений - секретов, которые используются в других частях организма. Для того чтобы транспортный пузырек с секретом слился с цитолеммой, необходимы сигналы извне. Такой экзоцитоз называют регулируемым. Сигнальные молекулы, способствующие выведению секретов, называют либеринами (рилизинг- факторами), а препятствующие выведению - статинами. Мембрана транспортного пузырька встраивается в цитолемму и становится ее частью. Экзоцитоз и возврат мембран эндосом в нормально функционирующей клетке уравновешены с поглощением мембран в ходе пино- и фагоцитоза.
Внутриклеточный синтез.Управление внутриклеточным синтезом осуществляется из ядра клетки. На активных участках хромосом синтезируются молекулы РНК. Они транспортируются к поровым комплексам и поступают в цитоплазму. На рибосомах из аминокислот происходит сборка белков, которые в соответствии с их назначением можно отнести к трем группам. Одна группа - это структурные белки, которые используются клеткой для построения собственных органелл, другая - белки, выделяемые клеткой вовне, это ее секреты; третья группа - ферменты, которые обеспечивают все внутриклеточные биохимические превращения как катализаторы. Часть ферментов остается в цитоплазме. Одни из них функционируют в гиалоплазме, другие встраиваются в органеллы. Третья часть ферментов направляется в ядро и там регулирует считывание генетической информации с ДНК и матричный синтез РНК.
В ядро возвращаются те белки, которые участвуют в построении самих хромосом.
Вещества, которые синтезируются на мембранах эндоплазматической сети, поступают в транспортные пузырьки и доставляются к комплексу Гольджи. В нем протекают заключительные этапы синтеза.
|
|
|
Синтез белка (трансляция) связан с процессом транскрипции - переписывания информации, хранящейся в ДНК. Благодаря ядерной оболочке в клетках процессы транскрипции и трансляции проходят в разных структурах и разделены во времени.
Транскрипция осуществляется в ядре. Информация о структуре белка, заключенная в ДНК, «переписывается» наинформационную РНК (иРНК). При этом с одного гена может «переписываться» множество молекул иРНК, которые транспортируются из ядра в цитоплазму, где и выполняют свои функции.
В клетках существует 3 типа РНК. Среди них иРНК переносит информацию о нуклеотидной последовательности ДНК к рибосомам. В образовании рибосом участвует рибосомная РНК (рРНК). Небольшие транспортные РНК (тРНК) выполняют двойную функцию: они присоединяют молекулу аминокислоты, транспортируют ее к рибосоме и узнают триплет, соответствующий этой аминокислоте в молекуле иРНК. Рибосомная и транспортная РНК (рРНК и тРНК) синтезируются на идентичных генах, которые (в отличие от генов иРНК) в каждой клетке имеются в виде множества копий. Реакции синтеза белка осуществляют рибосомы, которые считывают информацию. Перемещаясь по цепи иРНК, рибосома присоединяет аминокислоты, которые связываются между собой, а молекулы тРНК отделяются, чтобы вскоре присоединить новую аминокислоту.
Таким образом, следует отметить что клеточные процессы строго зависимы друг от друга и протекают последовательно один за одним выполняя все стадии жизненного цикла клетки – обмен веществ, раздражение, движение, размножение.
Основные процессы клеточного метаболизма
В нормально функционирующей живой клетке одномоментно протекает множество катализируемых ферментами химических реакций, приводящих к образованию огромного количества разнообразных соединений. В норме обмен веществ в клетке (метаболизм) осуществляется по принципам строжайшей экономии энергии и вещества, что обеспечивается сложнейшей системой регуляции обмена веществ.
Метаболизм – это химические реакции, возникающие с момента поступления в организм питательных веществ до момента выделения во внешнюю среду конечных продуктов этих реакций. Это сложный процесс преобразования потребляемой пищи в жизненную энергию. В метаболизм вовлечены все реакции, протекающие в живых клетках, результатом которых является строительство структур тканей и клеток. То есть, метаболизм можно рассматривать, как процесс обмена в организме веществ и энергии.
Живая клетка представляет собой высокоорганизованную систему, включающую различные структуры, а также специальные ферменты, способные эти структуры разрушить. Содержащиеся в клетке макромолекулы путём гидролиза могут распадаться на мелкие составляющие. В клетке обычно очень мало натрия и много калия, при этом она существует в среде, где мало калия и много натрия, причём проницаемость клеточной мембраны для обоих ионов одинакова. Отсюда вывод: клетка – это весьма далёкая от химического равновесия система.
Для поддержания клетки в химически неуравновешенном состоянии организму требуется производить определённую работу, для которой необходима энергия. Получение энергии для выполнения этой работы является непременным условием того, чтобы клетка пребывала в своём нормальном стационарном химически неуравновешенном состоянии. Одновременно в клетках выполняется и другая работа по взаимодействию со средой, к примеру: проведение нервных импульсов в нервных клетках, сокращение мышц – в мышечных, образование мочи в клетках почек и прочее.
Питательные вещества, попав внутрь клетки, начинают метаболизироваться, или претерпевать множество химических изменений и образовывать промежуточные продукты – метаболиты. Метаболический процесс в целом подразделяется на две категории: анаболизм и катаболизм. При анаболических реакциях из простых молекул путём биосинтеза образуются сложные молекулы, что сопровождается затратой свободной энергии. Анаболические превращения обычно восстановительные. При катаболических реакциях, наоборот, поступившие с пищей и входящие в состав клетки сложные компоненты расщепляются до простых молекул. Эти реакции преимущественно окислительные, сопровождающиеся выделением свободной энергии.
Непосредственным источником энергии, используемой клеткой для производства работы, служит энергия, заключённая в молекуле аденозинтрифосфата (АТФ).

Рисунок 3 – Молекула аденозинтрифосфата
В силу некоторых своих структурных особенностей, соединение АТФ богато энергией, и происходящий в ходе метаболического процесса разрыв связей фосфатных групп осуществляется таким образом, что высвободившаяся энергия может быть использована. Однако, в результате простого гидролиза разрыв фосфатных связей молекулы АТФ сделает высвобождённую для клетки энергию недоступной, поскольку метаболический процесс должен последовательно состоять из двух этапов с участием в каждом из них промежуточного продукта, в противном случае энергия выделяется в виде тепла и расходуется впустую. Молекула АТФ необходима практически для всех проявлений жизнедеятельности клеток, поэтому, неудивительно, что активность живых клеток в первую очередь направлена на синтез АТФ. Этот процесс состоит из сложных последовательных реакций с использованием потенциальной химической энергии, заключённой в молекулах жиров и углеводов.
Анаболизм тесно связан с катаболизмом, поскольку из продуктов распада питательных веществ получаются новые вещества. Если анаболизм направлен на образование составных структур клеток и тканей, то катаболизм превращает сложные молекулы в простые. Простые молекулы частично используются на биосинтез (образование органических веществ из простых соединений под действием ферментов-биокатализаторов), и частично выводятся из организма в виде продуктов распада, таких как мочевина, аммиак, диоксид углерода и вода.
Между катаболитическими и анаболитическими процессами в клетке существует тесная взаимосвязь. Катаболитические процессы служат источником энергии и “строительного материала” для анаболитических процессов, а продукты анаболизма могут служить субстратом для катаболитичких процессов (питательные вещества) или выполнять функции катализаторов (белки-ферменты).
Самый простой способ регуляции любого метаболического пути основывается на доступности субстрата. Действительно, в соответствии с законом действия масс, снижение количества субстрата-реагента (его концентрации в среде) приводит к снижению скорости протекания процесса (реакции) через данный метаболический путь. С другой стороны, повышение концентрации субстрата приводит к стимулированию этого метаболического пути. Поэтому, независимо от каких-то иных факторов, наличие (доступность) субстрата является важнейшим механизмом интенсификации любого метаболического процесса. Иногда эффективным средством повышения выхода целевого продукта является увеличение концентрации в клетке какого-либо определенного предшественника. Однако, в отличие от химических процессов, в биотехнологии данный путь имеет свои ограничения, т.к. высокие концентрации субстратов (больше 3-5%), например глюкозы или сахарозы, обычно резко тормозят рост микроорганизмов, что используется, например, для консервирования ягод и фруктов. Связано это, прежде всего с осмотическим эффектом, который вызывается большой разностью в концентрации этих веществ внутри клеток и в окружающей среде.
Однако в клетках имеется на много порядков более эффективный механизм контроля метаболитических процессов, основанный на регуляции ферментативного аппарата клетки. Такая регуляция может осуществляться по крайней мере двумя путями. Один из них очень быстрый (реализующийся в течение секунд или минут) заключается в изменении каталитической активности уже имеющихся молекул фермента. Второй, более медленный (реализуется в течение многих минут), состоит в изменении скоростей синтеза (количества) ферментов. В обоих механизмах используется единый принцип управления системами – принцип обратной связи.
Поскольку все процессы протекающие в клетке требуют участия специфических белковых катализаторов – ферментов, то общее количество ферментов в клетках может варьироваться от нескольких десятков до нескольких сотен, а процентная доля их по отношению к другим клеточным белкам будет достаточно большой (до нескольких процентов даже для одного фермента).
Однако энергетических (АТФ) и сырьевых ресурсов клетки (аминокислот) не хватает для одновременного синтеза всех необходимых ферментов. Поэтому постоянно синтезируются только те ферменты, которые поддерживают основные клеточные функции (например ферменты гликолиза, ЦТК). Такие ферменты называют конститутивными. Другие ферменты, адаптивные или индуцибельные, синтезируются только в ответ на появление каких то внешних факторов или веществ – индукторов, которые являются субстратами (питательными веществами) или их аналогами.
Уровень синтеза таких ферментов регулируется двумя механизмами – индукцией и репрессией.
Под индукцией понимают относительное увеличение синтеза одного фермента или группы ферментов, участвующее в одной и той же последова-тельности реакций, например в разложении какого-то сложного вещества до более простых. Ферменты, синтез которых регулируется таким образом, называют адаптивными или индуцированными (индуцибельными), а субстраты, вызывающие их синтез - индукторами. Под влиянием индукторов количество адаптивных ферментов может возрастать в сотни раз. Так, для E.coli установлено, что у культуры, выросшей на среде с глюкозой, обнаруживает лишь следы β-галактозидазы, осуществляющей реакцию расщепления лактозы до α -галактозы и D-глюкозы. При перенесении культуры на среду с лактозой, уже через несколько минут, начинается активный синтез β-галактозидазы и у адаптированной культуры до 3% от содержания белка приходится на этот фермент.
Для индуцируемых ферментов установлено, что:
а) фермент появляется во всех клетках одновременно и это нельзя объяснить мутациями;
б) индуцированный фермент целиком синтезируется в клетке из аминокислот или, как говорят, образуется de novo (изначально).
в) фермент синтезируется до тех пор, пока в среде есть индуктор. Через индукцию регулируется синтез ферментов, участвующих в катаболических процессах, т.е. индуцируемые ферменты необходимы для поглощения клеткой субстратов и включения их в обмен.
При промышленном получении ферментов, часто великолепными индукторами являются неутилизируемые структурные аналоги субстратов. Например, для β-галактозидазы таким веществом служит изопропил-β – D-тио-галактопиранозид (ИПТГ) неметаболизируемый аналог лактозы. Это позволяет увеличить выход фермента, который при этом не расходуется в ферментативной реакции и облегчить его очистку т.к. ИПТГ берется в количестве значительно меньшем, чем лактоза и в культуральной жидкости нет продуктов ее распада.
Вторым механизмом регуляции синтеза ферментов является репрессия, когда наблюдается относительное уменьшение синтеза фермента или группы ферментов, участвующих в одной и той же последовательности реакций, В зависимости от природы репрессоров различают репрессию конечным продуктом и репрессию катаболитами. Репрессия конечным продуктом наблюдается только для ферментов, осуществлявших анаболические реакции. При наличии в клетке конечного продукта анаболического пути снижается скорость синтеза всех ферментов, участвующих в его образовании. Этот процесс позволяет экономить клеточный белок, останавливая синтез тех ферментов, которые в данный момент не требуются клетке.
Репрессия катаболитами характерна для реакций разложения сложных органических веществ микроорганизмами. Этот механизм позволяет клетке использовать более доступный субстрат, обеспечивавший высокую скорость роста культуры. Предпочтение отдается тем субстратам, разложение которых включает меньшее число стадий: микроорганизмы предпочитают простые сахара сложным, аминокислоты - пептидам и т.д. Одним из примеров катаболитной репрессии является “глюкозный эффект" - явление, наблюдаемое при выращи-вании микроорганизмов на средах, содержащих наряду с глюкозой другие источники углерода. Глюкоза, как наиболее легко усвояемый субстрат, метаболизируется в клетке и продукты ее разложения тормозят синтез ферментов, участвующих в усвоении более сложных субстратов до тех пор, пока не будет использована вся глюкоза.
Регуляция обмена веществ микробной клетки может происходить также путем изменения ферментативной активности имеющихся ферментов. Это явление наблюдается преимущественно в анаболитических процессах. Наиболее изученным механизмом является ингибирование активности ферментов конечным продуктом (ретроингибирование), когда активность фермента, стоящего в начале многоступенчатого превращения субстрата тормозится конечным метаболитом.
Впервые о наличии такого регуляторного механизма было сообщено в 1953 г. При изучении биосинтеза триптофана клетками E.coli. Заключительный этап биосинтеза данной ароматической аминокислоты состоит из нескольких, катализируемых индивидуальными ферментами стадий. Было обнаружено, что у одного из мутантов E. coli с нарушенным биосинтезом триптофана добавление данной аминокислоты (являющейся конечным продуктом этого биосинтетического пути) резко тормозит накопление одного из предшественников – индол глицерофосфата в клетках. Уже тогда было высказано предположение, что триптофан ингибирует активность какого-то фермента, катализирующего образование индол глицерофосфата. Несколько позднее было четко установлено, что таким чувствительным к триптофану ферментом является антранилатсинтетаза, которая катализирует более раннюю реакцию триптофанового пути – образование антраниловой кислоты из хоризмовой кислоты и глутамина. Этот факт был экспериментально обоснован в опыте, когда добавление триптофана в клеточные экстракты E. coli, содержащие фермент антранилатсинтетазу и его субстраты (хоризмат и глутамин), приводило к резкому ингибированию образования антранилата. Более того, было однозначно продемонстрировано, что активность антранилатсинтетазы подавляется только триптофаном и никакие другие метаболиты клетки подобного действия не оказывают.
Благодаря этому явлению у микроорганизмов предотвращается перепроизводство низкомолекулярных промежуточных продуктов обмена, таких, как аминокислоты, пуриновые и пиримидиновые нуклеотиды. Как правило, субстрат ингибируемого фермента резко отличается от конечного продукта - ингибитора и это обстоятельство позволяет считать, что конечный продукт соединяется не с активным центром фермента, а со специальным регуляторным или аллостерическим (от греч. «аллос» – другой, «стерос» – пространственный), центром. Присоединение конечного продукта к аллостерическому центру фермента сопровождается утратой нормальной каталитической активности вследствии конформационных изменений структуры белковой молекулы.
По сравнению с индукцией и репрессией ретроингибирование это инструмент быстрого и точного регулирования метаболитических процессов.
Ретроингибирование является крайне нежелательным явлением при промышленном получении тех или иных интересующих человека клеточных метаболитов, т.к. препятствует их накоплению в высоких концентрациях, что требует использования установок большего объема и усложняет процесс их выделения и очистки. А это в свою очередь увеличивает себестоимость продукции. Существует несколько подходов, позволяющих снять или значительно уменьшить эффект ретроингибирования. Один из них состоит в том, что целевой продукт (ингибитор), удаляют. Например, если он является эндометаболитом, то создаются условия для его ухода из клетки в культуральную жидкость, например за счет повышения проницаемости клеточных оболочек. Если целевой продукт является экзометаболитом (аминокислоты, антибиотики), то его удаляют из культуральной жидкости, например, переводя в нерастворимое состояние (осадок). Второй подход состоит в том, что на стадии синтеза продукта в культуральную жидкость добавляют вещество-промежуточный метаболит, синтез которого блокируется конечным продуктом (см. синтез триптофана). Недостатком этого подхода является то, что такой предшественник не всегда может быть получен дешево и в больших количествах. На практике, если возможно, обычно применяют оба подхода.
Другие подходы связаны с использованием методов мутагенеза-селекции и генной инженерии. Например, при мутационном изменении аллостерического центра (центра взаимодействия с ингибитором) чувствительность к ингибитору утрачивается и фермент сохраняет свою активность при высоких концентрациях конечного продукта, что позволяет создать более высокопродуктивные штаммы микроорганизмов-продуцентов. Более сложный вариант данного подхода реализуется при микробиологическом получении лизина.
Таким образом, клеточный метаболизм выполняет четыре основные специфические функции: извлечение энергии из окружающей среды и преобразование ее в энергию макроэргических (высокоэргических) соединений в количестве, достаточном для обеспечения всех энергетических потребностей клетки; образование из экзогенных веществ (или получение в готовом виде) промежуточных соединений, являющихся предшественниками высокомолекулярных компонентов клетки; синтез белков, нуклеиновых кислот, углеводов, липидов и других клеточных компонентов из этих предшественников; синтез и разрушение специальных биомолекул, образование и распад которых связаны с выполнением специфических функций данной клетки. Для понимания сущности обмена веществ и энергии в живой клетке нужно учитывать ее энергетическое своеобразие. Все части клетки имеют примерно одинаковую температуру, т. е. клетка изотермична. Различные части клетки мало отличаются и по давлению. Это значит, что клетки не способны использовать в качестве источника энергии тепло, т. к. при постоянном давлении работа может совершаться лишь при переходе тепла от более нагретой зоны к менее нагретой, таким образом, живая клетка представляет собой изотермическую химическую машину.
Заключение
Таким образом, следует отметить что клеточные процессы строго зависимы друг от друга и протекают последовательно один за одним выполняя все стадии жизненного цикла клетки – обмен веществ, раздражение, движение, размножение.
Из приведенного определения клетки становится ясно, что клетка представляет собой ограниченную активной мембраной, упорядоченную структурированную систему биополимеров (белков, нуклеиновых кислот) и их макромолекулярных комплексов, участвующих в единой совокупности метаболических и энергетических процессов, осуществляющих поддержание и воспроизведение всей системы в целом.
Клеточный метаболизм выполняет четыре основные специфические функции: извлечение энергии из окружающей среды и преобразование ее в энергию макроэргических (высокоэргических) соединений в количестве, достаточном для обеспечения всех энергетических потребностей клетки; образование из экзогенных веществ (или получение в готовом виде) промежуточных соединений, являющихся предшественниками высокомолекулярных компонентов клетки; синтез белков, нуклеиновых кислот, углеводов, липидов и других клеточных компонентов из этих предшественников; синтез и разрушение специальных биомолекул, образование и распад которых связаны с выполнением специфических функций данной клетки.
Таким образом, метаболические процессы, проходящие в клетке, тесно связаны с процессами всей клетки и являются основной частью клеточной системы в целом, т.к. метаболические процессы есть совокупность процессов превращения веществ и энергии в живом организме и обмена организма веществами и энергией с окружающей средой.
|
|
|
