 |


Отбор как избирательное воспроизведение генотипов
|
|
|
|
Ч. Дарвин определил естественный отбор как сохранение особей с полезными и гибель с вредными индивидуальными уклонениями – «переживание наиболее приспособленных и гибель менее приспособленных». Однако, в процессе естественного отбора важны не столько выживание или гибель особей, а вклад каждой особи в генофонд популяции. Без размножения вклад особей в генофонд популяций равен нулю. Только успех в распространении и закреплении определенных аллелей (или целых генных комплексов) в популяциях ведет к возникновению элементарного эволюционного явления, лежащего в основе всякого эволюционного процесса, но достигается это лишь при дифференциальном воспроизведении генотипов. Успех в размножении разных особей может служить объективным генетико-эволюционным критерием естественного отбора. Следовательно, под естественным отбором нужно понимать избирательное (дифференциальное) воспроизведение генотипов или генных комплексов (А.В. Яблоков, А.Г. Юсуфов, 2006).
Естественный отбор как движущий фактор эволюции имеет вероятностный закономерный характер; формируя признаки и свойства путем интегрирования отдельных случайных уклонений с незначительной адаптивной ценностью, он обладает накапливающим и интегрирующим действием, ему свойственна адаптивная направленность. Направленность в действии отбора возникает стихийно. Естественный отбор никогда не ведет к закреплению признаков, вредных для вида, он не предопределен, не направлен заранее к какой-то цели. Направление отбора определяется соотношением наследственной изменчивости в генофонде популяции и условий борьбы за существование. Если признак оказывается более адаптивным при данных условиях по сравнению со всеми уклонениями от средней нормы по этому признаку, отбор будет сохранять такой вариант, а все уклоняющиеся варианты будут элиминироваться. В альтернативной ситуации, если средняя норма признака оказывается малоадаптивной в данных условиях, отбор будет подхватывать любые уклонения от нормы, то есть его направление будет противоположным (А.С. Северцов, 1987, 2005).
|
|
|
Естественный отбор может действовать по любому признаку или свойству, изменяя любой наследственный признак в череде поколений. Естественный отбор на каждом из этапов эволюции оставляет особи с особенностями, лучшими в определенных условиях. Если условия существования группы сохраняются стабильными достаточно длительное время, направление действия отбора в ряду поколений также остается стабильным. Поэтому можно признать, что именно отбор направляет эволюцию по определенному руслу. Направление для естественного отбора определяется сложным сочетанием внешних и внутренних условий существования, успехом в борьбе за существование.
Механизм действия естественного отбора заключается в сохранении, накоплении и усилении в ряду поколений мелких, индивидуальных, наследственных уклонений в признаках и свойствах (накоплении и усилении в ряду поколений случайных уклонений с незначительной адаптивной ценностью). Под отбор попадают индивиды, группы – семьи, популяции, группы популяций, виды, целые сообщества. Первично отбор действует в популяциях – это поле действия отбора. В пределах популяций отбираются индивиды, обладающие какими-то достоинствами перед другими. Эти достоинства выражены в наличии или отсутствии каких-либо признаков и свойств, рассматриваемых как точки приложения отбора. Но борьба за существование происходит не между признаками и генами, их определяющими, а между их носителями – особями. Именно индивиды являются элементарными объектами отбора. Сфера действия естественного отбора затрагивает жизненно важные признаки и свойства особи. Отбор всегда идет по фенотипам. В фенотипе особи отражаются особенности генотипа, и в ряду поколений отбор по фенотипам сводится к отбору определенных генотипов. В итоге, сферой действия отбора являются фенотип и генотип. Двойственность и ступенчатость в действии отбора определяет важность фенотипа и фенотипической изменчивости в эволюции.
|
|
|
коэффициент отбора (S) – это показатель, который определяет приспособленность организма. Приспособленность организма оценивается по выживаемости – чем выше его выживаемость, тем выше приспособленность. Степень относительной приспособленности генотипа (W) характеризуется его адаптивной ценностью – способностью генотипа к выживанию и воспроизведению по сравнению с другими генотипами в популяциях. Коэффициент отбора варьирует от 1 до 0 и представляет собой величину, обратную адаптивной ценности генотипа. Чем больше адаптивная ценность генотипа, тем ниже коэффициент отбора. Если W = 1, то S = 0. Например, из 100 родиишихся особей с определенным признаком до размножения доживут все, а с другим признаком – лишь 90 особей. Это значит, что в первом случае коэффициент отбора равен 0, а во втором – 0,1. В природных условиях коэффициент отбора обычно не превышает 0,10 - 0,20, чаще он имеет меньшие значения.
Коэффициент отбора может выражать как возрастание, так и убывание концентраций аллеля. В первом случае имеет место размножение носителя, во втором – его элиминация.
Эффективность отбора – под ней понимают степень достижения к определенному моменту времени положительного результата отбора, который определяется совокупностью положительно отбираемых генотипов и связанных с ними морфо-физиологических свойств организмов. Эффективность отбора определяется по формуле: 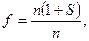 то есть она определяется отношением чисел выживающих особей каждого варианта. Если, например, она равна 1,5, то это означает, что один из сравниваемых вариантов в 1,5 раза лучше приспособлен по сравнению с другим.
то есть она определяется отношением чисел выживающих особей каждого варианта. Если, например, она равна 1,5, то это означает, что один из сравниваемых вариантов в 1,5 раза лучше приспособлен по сравнению с другим.
Скорость отбора – этот параметр определяется величиной приращения отбираемого варианта в единицу времени. Она пропорциональна коэффициенту отбора и выражается формулой: 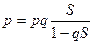 , где р – концентрация отбираемого варианта, q – концентрация элиминируемого варианта. Имея значения коэффициента отбора и концентрацию отбираемого варианта, можно установить скорость отбора по данному варианту. Описанная модель скорости отбора по двум альтернативным вариантам является упрощенной моделью. Уже при трех вариантах, учитывающих три генотипических класса, применяются другие формулы. На скорость отбора влияют ряд факторов. Возможность естественного отбора обусловливается наличием материала для отбора, то есть генетическим многообразием особей данной популяции. Возникновение неблагоприятных мутаций замедляет его, возникновение благоприятных мутаций ускоряет течение отбора. Максимально возможная скорость естественного отбора определяется масштабом изменчивости в признаках, имеющих какое-либо значение в борьбе за существование. В математическом выражении это положение известно как закон Р. Фишера. По данному закону скорость отбора признака пропорциональна квадрату среднего квадратического уклонения данного признака. Из закона Р. Фишера следует, что отбор не может снижать приспособленность. Все особи, обладающие относительной приспособленностью более низкой, чем средняя приспособленность популяции, как правило, гибнут. Поэтому наиболее очевидный эффект естественного отбора – это поддержание некоего уровня приспособленности особей популяции, позволяющего ей существовать в данных условиях среды. Средняя приспособленность популяции изменяется под действием отбора тем быстрее, чем ниже начальное значение приспособленности и чем выше ее изменчивость. Р. Фишер математически показал, что направленное преобразование генофонда, то есть эволюция, идет наиболее эффективно в том случае, если популяции изолированы не полностью и обмениваются генами через ограниченное, но все же существующее скрещивание. В этом случае приток аллелей извне постоянно поддерживает генетическое разнообразие особей популяции, а изоляция сохраняет своеобразие ее генофонда и тем самым своеобразие вырабатываемых адаптаций. Скорость естественного отбора зависит не только от изменчивости популяции, то есть резерва накопленных изменений, но и от скорости мутирования. Скорость мутирования обычно незначительна, но отбору подлежат не отдельные мутации, а сложные их комбинации, число которых значительно больше числа мутаций. Поэтому в природных популяциях всегда имеется материал для отбора. На скорость протекания естественного отбора влияют размеры популяций. Благоприятны для быстрого действия естественного отбора популяции с колеблющейся численностью (что было отмечено выше).
, где р – концентрация отбираемого варианта, q – концентрация элиминируемого варианта. Имея значения коэффициента отбора и концентрацию отбираемого варианта, можно установить скорость отбора по данному варианту. Описанная модель скорости отбора по двум альтернативным вариантам является упрощенной моделью. Уже при трех вариантах, учитывающих три генотипических класса, применяются другие формулы. На скорость отбора влияют ряд факторов. Возможность естественного отбора обусловливается наличием материала для отбора, то есть генетическим многообразием особей данной популяции. Возникновение неблагоприятных мутаций замедляет его, возникновение благоприятных мутаций ускоряет течение отбора. Максимально возможная скорость естественного отбора определяется масштабом изменчивости в признаках, имеющих какое-либо значение в борьбе за существование. В математическом выражении это положение известно как закон Р. Фишера. По данному закону скорость отбора признака пропорциональна квадрату среднего квадратического уклонения данного признака. Из закона Р. Фишера следует, что отбор не может снижать приспособленность. Все особи, обладающие относительной приспособленностью более низкой, чем средняя приспособленность популяции, как правило, гибнут. Поэтому наиболее очевидный эффект естественного отбора – это поддержание некоего уровня приспособленности особей популяции, позволяющего ей существовать в данных условиях среды. Средняя приспособленность популяции изменяется под действием отбора тем быстрее, чем ниже начальное значение приспособленности и чем выше ее изменчивость. Р. Фишер математически показал, что направленное преобразование генофонда, то есть эволюция, идет наиболее эффективно в том случае, если популяции изолированы не полностью и обмениваются генами через ограниченное, но все же существующее скрещивание. В этом случае приток аллелей извне постоянно поддерживает генетическое разнообразие особей популяции, а изоляция сохраняет своеобразие ее генофонда и тем самым своеобразие вырабатываемых адаптаций. Скорость естественного отбора зависит не только от изменчивости популяции, то есть резерва накопленных изменений, но и от скорости мутирования. Скорость мутирования обычно незначительна, но отбору подлежат не отдельные мутации, а сложные их комбинации, число которых значительно больше числа мутаций. Поэтому в природных популяциях всегда имеется материал для отбора. На скорость протекания естественного отбора влияют размеры популяций. Благоприятны для быстрого действия естественного отбора популяции с колеблющейся численностью (что было отмечено выше).
|
|
|
|
|
|
Доказательства действия естественного отбора. В ХIХ - ХХ вв. в развитых индустриальных районах происходило потемнение окраски у многих бабочек – темная окраска оказалась покровительственной в промышленных загрязненных районах. Наблюдения показали, что некоторые насекомоядные птицы в промышленных центрах вылавливают в основном светлых бабочек, а вдали от них – темных бабочек. В Европе в настоящее время более чем у 70 видов чешуекрылых окраска изменилась. Это показывает на существование сильного отбора, направленного против темных форм в незагрязненных местностях и против светлых в загрязненных районах. Распространение меланической формы определяется поддержкой естественным отбором доминантного гена, ответственного за темную окраску и связанного с изменением поведения (бабочки активно выбирали темный фон субстрата и были менее подвижны). Изучены микроэволюционные изменения в популяциях ужей, обитающих на островах оз. Эри в Северной Америке: на белых известняковых скалах островов выживают преимущественно светлые особи, полосатая окраска оказывается защитной в болотистых местах. Под действием естественного отбора происходит возникновение внутрипопуляционного полиморфизма в популяциях. Примером такового является распространение серповидноклеточной анемии в некоторых тропических районах. Естественный отбор, направленный на выживание и размножение особей, покровительствует индивидуумам, гетерозиготным по гену серповидноклеточности (ген серповидноклеточности определяет устойчивость особи к малярии). Много примеров ведущего действия отбора описано в связи с повышением резистентности (устойчивости) микроорганизмов – к антибиотикам, насекомых и крыс к ядохимикатам. Так, в Великобритании при использовании в 50-е годы ХХ в. ядохимиката варфарина для борьбы с крысами сначала были получены хорошие результаты – крысы почти полностью исчезали на значительной территории, но некоторые особи из миллионов крыс случайно обладали наследственным иммунитетом против варфарина, для них поедание приманки не было смертельным; от этих немногих особей возникли популяции устойчивые к действию варфарина. К 1972 г. вся страна была вновь населена крысами (рисунок 7).
Признание за естественным отбором ведущей роли в эволюции не принижает значения остальных факторов эволюционного процесса. Все элементарные эволюционные факторы взаимосвязаны под контролем отбора, все они воздействуют на элементарный эволюционный материал, изменяя элементарную эволюционную единицу.
|
|
|
