 |


Закономерности воздействия хищников на популяцию
|
|
|
|
Потребление рыб другими организмами, в том числе рыбами, является одной из важнейших причин смертности. У каждого вида рыбы, особенно на ранних этапах онтогенеза, хищники составляют обычно один из важнейших элементов среды, приспособления к которому бывают весьма разнообразны. Большая плодовитость рыб, охрана потомства, покровительственная окраска, различные защитные приспособления (шипы, колючки, ядовитость и др.), защитные особенности поведения представляют собой различные формы приспособлений, обеспечивающие существование вида в условиях определенного пресса хищников.
В природе нет видов рыб, которые были бы свободны от большего или меньшего, но закономерного воздействия хищников. Одни виды подвержены этому воздействию в большей степени и на всех этапах онтогенеза, например анчоусы, особенно мелкие, сельдевые, бычки и пр. Другие подвергаются этому воздействию в меньшей степени и главным образом на ранних этапах развития. На более поздних этапах развития у некоторых видов воздействие хищников может сильно ослабевать и практически сходить на нет. К этой группе рыб относятся осетровые, крупные сомы, некоторые виды карповых. Наконец, третья группа - это виды, у которых смерть от хищников и на ранних этапах онтогенеза очень невелика. К этой группе относятся только некоторые акулы и скаты. Естественно, что границы между этими выделенными нами группами условны. У рыб, приспособленных к значительному прессу хищников, меньший процент умирает от старости в результате старческого нарушения обмена веществ.
Большая или меньшая защищенность от хищников соответственно связана с выработкой способности компенсировать большую или меньшую гибель путем изменения темпа воспроизводства популяции. Виды, приспособленные к значительному выеданию хищниками, могут и компенсировать большие потери. Приспособление к определенному характеру воздействия хищников формируется у рыб, как и у других организмов, в процессе формирования фаунистического комплекса. В процессе видообразования происходит коадаптация хищника и жертвы. Виды-хищники приспосабливаются питаться определенного типа жертвами, а виды-жертвы приспосабливаются теми или иными способами ограничивать воздействие хищников и компенсировать убыль.
|
|
|
Хищниками в широком смысле, включая и паразитизм, по отношению к рыбам являются почти все группы организмов от растений и простейших до млекопитающих. Но основными хищниками рыб являются сами же рыбы, некоторые группы беспозвоночных (кишечнополостные, моллюски, преимущественно головоногие, ракообразные и насекомые) - главным образом в отношении ранних этапов онтогенеза. Интенсивность воздействия хищников и выработка в связи с этим соответствующих защитных и компенсаторных приспособлений оказываются различными в водоемах разного типа и разных широт.
Выше мы рассмотрели закономерности изменения плодовитости и, в частности, показали, что популяции одних и тех же видов в низких широтах оказываются плодовитее, чем в высоких. Близкие формы Тихого океана оказываются плодовитее, чем формы Атлантики. Рыбы рек Дальнего Востока плодовитее, чем рыбы рек Европы и Сибири. Эти различия в плодовитости связаны с разным прессом хищников в указанных водоемах. Защитные приспособления вырабатываются у рыб применительно к жизни в соответствующих местах обитания. У рыб пелагиали основные формы защиты - это соответствующая "пелагическая" защитная окраска, быстрота движения и - для защиты от так называемых дневных хищников, ориентирующихся при помощи органов зрения, - стаеобразование. Защитное значение стаи, видимо, трояко. С одной стороны, рыбы в стае обнаруживают хищника на более далеком расстоянии и могут от него скрыться (Никольский, 1955). С другой стороны, стая представляет и определенную физическую защиту от хищников (Мантейфель и Радаков, 1960, 1961). Наконец, как это отмечено в отношении трески (хищник) и молоди сайды (жертва), множественность жертв и защитные маневры стаи дезориентируют хищника и затрудняют ему поимку добычи (Радаков, 1958, 1972; Hobson, 1968).
|
|
|
Защитное значение стаи сохраняется у многих видов рыб не на всех этапах онтогенеза. Обычно оно свойственно ранним этапам: у взрослых рыб стайный образ жизни, теряющий защитную функцию, проявляется лишь в отдельные периоды жизни (нерест, миграция). Стая как защитное приспособление свойственна обычно молоди рыб во всех биотопах как в пелагиали, так и в прибрежной зоне морей, как в реках, так и в озерах. Стая служит защитой от дневных хищников, но облегчает отыскание рыб в стае ночным хищникам, ориентирующимся в поиске пищи при помощи других органов чувств. Поэтому у многих рыб, например у сельдевых, на ночь стая распадается и особи держатся в одиночку, чтобы с рассветом вновь собраться в стаю.
У прибрежных придонных и донных рыб различны и способы защиты от хищников. Основную роль приобретают различные морфологические защитные приспособления, различные шипы и колючки.
Развитие "вооружений" у рыб против хищников далеко не одинаково в разных фаунах. В фаунах морей и пресных вод низких широт "вооруженность" развита обычно интенсивнее, чем в фаунах более высоких широт (табл. 76). В фаунах низких широт относительное и абсолютное количество рыб, "вооруженных" шипами и колючками, гораздо больше, и "вооружение" у них развито сильнее. В низких широтах и ядовитых рыб больше, чем в высоких широтах. У морских рыб защитные приспособления в одних и тех же широтах развиты больше, чем у рыб пресных вод.

Таблица 76. Количество рыб с шипами и колючками в разных морских фаунах (без миног и хрящевых), %
Среди представителей древнеглубоководной фауны процент "вооруженных" рыб несравненно меньше, чем в фаунах континентального шельфа.
|
|
|
В прибрежной зоне "вооруженность" рыб развита гораздо сильнее, чем в открытой части моря. По побережью Африки, в районе Дакара в прибрежной зоне "вооруженные" виды рыб в траловых уловах составляют 67%, а вдали от берега их количество снижается до 44%. Несколько иная картина наблюдается в районе Гвинейского залива. Здесь в прибрежной зоне процент "вооруженных" видов очень невелик (только сомы Ariidae), а в отдалении от берега он значительно возрастает (Радаков, 1962; Radakov, 1963). Меньший процент "вооруженных" рыб в прибрежной зоне Гвинейского залива связан с высокой мутностью прибрежных вод этого района и невозможностью из-за этого охоты здесь "зрительным хищникам", которые концентрируются в прилегающих участках с прозрачной водой. В зоне с мутной водой менее многочисленные хищники представлены видами, ориентирующимися на жертву при помощи других органов чувств (см. ниже).
В морях Дальнего Востока аналогичное положение. Так, в Охотском море среди прибрежной зоны "вооруженных" рыб больше, чем вдали от берега (Шмидт, 1950). То же наблюдается и по американскому побережью Тихого океана.
Относительное количество "вооруженных" рыб различно также в северной части Атлантического океана и в Тихом океане (Clements a. Wilby, 1961): в северной части Тихого океана процент "вооруженных" рыб много выше, чем в Северной Атлантике. Сходная закономерность наблюдается и в пресных водах. Так, в реках бассейна Ледовитого океана "вооруженных" рыб меньше, чем в бассейне Каспия и Аральского моря. Разная "вооруженность" свойственна и рыбам, населяющим разные биотопы. В направлении от верховья к низовью реки относительная численность "вооруженных" рыб обычно увеличивается. Это отмечено в реках разных типов и широт. Например, в среднем и нижнем течениях Амударьи рыб, имеющих шипы и колючки, около 50, а в верхнем течении - около 30%. В среднем и нижнем течениях Амура "вооруженных" видов более 50, а в верхнем менее 25% (Никольский, 1956а). Правда, из этого правила бывают исключения в реках, текущих с юга на север в северном полушарии.
|
|
|
Так, в р. Оби, например, не удается подметить заметной разницы в "вооруженности" рыб верховья и низовья. В низовье процент "вооруженных" видов становится даже несколько меньше.
Весьма существенно различается и интенсивность или, так сказать, мощность развития "вооружений" в разных зонах. Как показал И. А. Паракецов (1958), близкие виды Северной Атлантики обладают менее развитым "вооружением", чем виды Тихого океана. Это хорошо прослеживается на представителях сем. Scorpaenidae и Cottidae (рис. 53).

Рис. 53. Связь роста преоперкулярного шипа (S) с изменением длины тела рыбы (L) у представителей Myoxocephalus (а) и Gymnacanthus (б) (по Паракецову, 1958): 1 - Myoxocephalus jaok Cuv. a. Val.; 2 - М. brandii Steind; 3 - M. scorpius L.; 4 - M. quadri-cornis L.; 5 - Gymnacanthus herzensteini Jord. et Staiks (Тихий океан); 6 - G. tricuspis Rnd. (Атлантика)
То же имеет место и в пределах разных зон Тихого океана. У более северных видов "вооружение" развито слабее, чем у их близких родственников, но распространенных южнее (Паракецов, 1962). У видов, распространенных на больших глубинах, спинные шипы развиты слабее, чем у родственных форм, распространенных в прибрежной зоне. Это хорошо показано у Scorpaenidae. Интересно, что в то же время, поскольку на глубинах относительные размеры жертв обычно крупнее (и иногда значительно), чем в прибрежной зоне, у глубинных "вооруженных" рыб обычно крупнее голова и сильнее развиты оперкулярные шипы (Phillips, 1961).
Таким образом, выявляется следующая общая закономерность: интенсивность воздействия хищников на популяции и соответственно развитие у рыб-жертв защитных приспособлений возрастают в направлении от высоких широт к низким и от верховий рек к низовьям. В общем, чем стабильнее абиотические условия той или иной зоны, тем выше обычно в этой зоне пресс хищников. Обратная картина наблюдается в направлении от глубин к прибрежной зоне океана: на глубинах абиотические условия оказываются стабильнее, чем в прибрежной зоне, однако и интенсивность воздействия хищников тоже, видимо, ниже. Соответственно и хищники низких широт оказываются приспособленными питаться лучше защищенной добычей, чем более высокоширотные хищники. Поэтому переход хищника более низких широт на питание более высокоширотными жертвами бывает обычно много легче, чем если хищники высоких широт переходят на питание жертвами более южного происхождения (Никольский, 1956а). Это относится также к паразитам. Более южные по своему происхождению паразиты обычно причиняют больший вред, нападая на северные жертвы, чем северные паразиты, нападая на жертвы более низких широт.
|
|
|
Естественно, что развитие шипов и колючек не создает абсолютной защиты от хищников, а лишь снижает интенсивность воздействия хищника на стадо жертвы. Как показали М. Н. Лишев (1950), И. А. Паракецов (1958), К. Р. Фортунатова (1959) и другие исследователи, наличие колючек делает рыб менее доступными для хищников, чем рыб сходного биологического типа и формы, но лишенных колючек. Наиболее наглядно это показано М. Н. Лишевым (1950) на примере поедания обыкновенного и колючего горчаков в Амуре. Защита от хищников обеспечивается не только наличием колючек (возможностью наколов), но и увеличением высоты тела, например у колюшки (Фортунатова, 1959), или ширины головы, например у подкаменщиков (Паракецов, 1958). Защитное значение шипов и колючек меняется в зависимости и от величины и способа охоты хищника, поедающего "вооруженную" рыбу, а также от поведения жертвы. Так, например, колюшка в дельте Волги оказывается доступной разным хищникам разных размеров. У окуня в пище встречаются самые мелкие рыбы, у щуки - более крупные и у сома - самые крупные (Фортунатова, 1959) (рис. 54). Как показано Фрост (Frost, 1954) на примере щуки, с увеличением размеров хищника возрастает и процент потребления им "вооруженных" рыб.
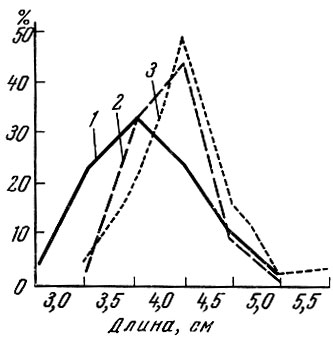
Рис. 54. Размеры половозрелой колюшки в пище окуня (1), щуки (2) и сома (3) (май-июль 1953 г.) (по Фортунатовой, 1959)
Интенсивность потребления "вооруженных" рыб в очень большой степени зависит и от того, насколько хищник обеспечен пищей. У голодных рыб при недостаточной кормовой базе интенсивность потребления "вооруженных" рыб возрастает. Это хорошо показано в эксперименте с колюшкой (Hoogland, Morris a. Tinbergen, 1956-1957). Здесь мы имеем частный случай общей закономерности, когда в условиях недостаточной обеспеченности основной, наиболее доступной пищей спектр питания расширяется за счет менее доступных кормов, на добычу и усвоение которых тратится больше энергии.
Существенное значение для доступности "вооруженных" рыб хищникам имеет поведение жертв. Как правило, рыбы выедаются хищниками в период их наибольшей активности. Это относится и к "вооруженным" рыбам. Например, девятииглая колюшка в дельте Волги доступнее всего хищникам в период размножения, в конце мая, и в период массового появления молоди, в конце июня - начале июля (рис. 55) (Фортунатова, 1959).

Рис. 55. Встречаемость колюшки в пище хищных рыб (май-июль 1953 г.), (по Фортунатовой, 1959)
Мы рассмотрели только две формы защиты жертвы от хищников: стайное поведение и "вооруженность" жертв, хотя формы защиты могут быть самыми разнообразными: это и использование тех или иных убежищ, например закапывание в грунт, и некоторые особенности поведения, например "крючок" у молоди сайды (Радаков, 1958), и вертикальные миграции (Мантейфель, 1961), и ядовитость мяса и икры, и многие другие способы. Интенсивность воздействия хищника на популяцию жертвы зависит от очень многих причин. Естественно, что каждый хищник приспособлен питаться в определенных условиях и определенными типами жертв. От характера местообитания жертв в очень большой степени зависит и специфика хищников, ими питающихся. В мутных водах рек Средней Азии основной тип хищников - это рыбы, ориентирующиеся на добычу при помощи органов осязания и органов боковой линии. Орган зрения у них в охоте за жертвами не играет существенной роли. Примерами могут служить большой лопатонос Pseudoscaphyrhynchus kaufmanni (Bogd.) и обыкновенный сом Silurus glanis L. Эти рыбы питаются и днем и ночью. В реках же с более прозрачной водой сом - типичный ночной хищник. В верхних течениях рек Европейского Севера и Сибири, где вода чистая и прозрачная, хищники (таймень Hucho taimen Pall., ленок Brachymystax lenok Pall., щука Esox lucius L.) ориентируются на добычу главным образом при помощи органа зрения и ведут охоту преимущественно в светлое время суток. В этой зоне только, пожалуй, налим Lota lota (L.), ориентирующийся на добычу главным образом при помощи обоняния, осязания и вкуса, кормится в основном ночью. То же наблюдается и в морях. Так, в прибрежных мутных водах Гвинейского залива хищники ориентируются в основном при помощи органов осязания и боковой линии. Орган зрения в этом биотопе играет у хищников подчиненную роль. Дальше от берега, за зоной мутной воды, в Гвинейском заливе в воде высокой прозрачности основное место занимают хищники, ориентирующиеся на добычу при помощи органа зрения, такие, как Sphyraena, Lutianus, тунцы и др. (Radakov, 1963).
Различны и способы охоты у хищников, добывающих пищу в зарослях и в открытых водах. В первом случае преобладают хищники-засадчики, во втором - ловящие добычу в угон. У многих хищников и в пределах одного места обитания четко выражена смена поедаемого корма в разное время суток: так, налим днем поедает малоподвижных беспозвоночных, а ночью охотится за рыбой (Павлов, 1959). Перкарина Perkarina maeotica Kuzn. в Азовском море днем питается главным образом веслоногими раками и мизидами, а ночью поедает тюльку Clupeonella delicatula Nordm. (Канаева, 1956).
Характер и интенсивность воздействия хищников на популяцию мирной рыбы зависят от многих причин: от абиотических условий, в которых осуществляется охота, от наличия и обилия других видов жертв, которыми питается тот же хищник; от наличия других хищников, питающихся той же жертвой; от состояния и поведения жертвы.
Резкие изменения абиотических условий могут очень сильно менять доступность жертвы для хищников. Так, например, в водохранилищах, где в результате значительных колебаний уровня исчезает подводная растительность, в прибрежной зоне резко ухудшаются условия охоты для хищника-засадчика щуки и, наоборот, создаются благоприятные условия для хищника более открытых вод - судака.
Каждый хищник приспособлен питаться определенным типом жертвы и, естественно, что наличие или отсутствие других видов жертв отражается на интенсивности их выедания. В этом отношении условия питания хищников изменяются особенно сильно, если появляются в массовом количестве жертвы, принадлежащие к другим, более северным фаунистическим комплексам. Так, например, в годы урожайные в Амуре на малоротую корюшку Hypomesus olidus (Pall.) весной, в период ее массового появления, все хищники переключаются на питание ею и, естественно, резко снижается их воздействие на других рыб (Лишев, 1950). Это наблюдалось, например, в 1947 г. и несколько в меньшей степени в 1948 г., а в неурожайный на корюшку 1946 г. хищники перешли на питание другой пищей и спектр питания их расширился.
Сходная картина наблюдается и в морях; так, в Баренцевом море в годы, урожайные на мойву, эта рыба весной составляет основу пищи трески. В случае же отсутствия или малого количества мойвы треска переходит на питание другими рыбами, в частности сельдью (Зацепин и Петрова, 1939).
Уменьшение численности жертвы, например молоди нерки в оз. Култус, приводит к тому, что обычно питающиеся ею хищники того же фаунистического комплекса переходят в значительной степени на питание другой, менее свойственной им добычей, перемещаясь при этом иногда в период нагула в менее обычные для них места обитания, где условия их питания оказываются хуже (Ricker, 1941).
Существенное влияние на интенсивность выедания хищником жертвы оказывает наличие другого хищника, поедающего ту же жертву, или наличие хищника, в отношении которого первый хищник является жертвой.
В случае охоты двух или нескольких хищников за одной жертвой доступность последней сильно возрастает. Это показано в эксперименте Д. В. Радаковым (1958), когда несколько хищников (треска) выедали жертв гораздо скорее, чем один хищник при той же плотности жертвы. Особенно повышается интенсивность выедания, если на рыбу одновременно охотятся хищники разных биологических типов. Одним из обычных способов защиты рыбы от хищника является выход в другое место обитания, где жертва оказывается недоступной для хищника, например уход от крупных хищников на мелководье или прижимание ко дну от пелагических хищников, или, наконец, выпрыгивание в воздух летучих рыб.
Если за жертвой охотятся одновременно хищники разных биологических типов (например, при скате молоди дальневосточных лососей гольцы Salvelinus и подкаменщики Myoxocephalus в речках, впадающих в лиман Амура), интенсивность выедания резко возрастает, ибо уход от пелагических хищников в придонные слои делает жертву доступнее для донных хищников и, наоборот, уход от дна в толщу воды повышает выедание пелагическими хищниками.
Часто весьма резко может меняться интенсивность выедания жертв хищниками, если последние сами находятся под воздействием хищника. Так, например, во время ската молоди горбуши и кеты из притоков Амура в низовьях притоков она в большом количестве выедается чебаком Leuciscus waleckii (Dyb.), причем, если здесь в низовье притока держится щука Esox reicherti Dyb., для которой чебак является основной пищей, активность чебака как потребителя скатывающейся молоди лососей резко снижается.
Сходная картина наблюдается в Черном море в отношении хамсы, ставриды и пеламиды. При отсутствии пеламиды Pelamys sarda (Bloch) ставрида Trachurus trachurus (L.) весьма интенсивно питается хамсой Engraulis encrassicholus L. В случае же появления пеламиды, для которой ставрида является жертвой, потребление ею хамсы резко снижается.
Естественно, что воздействие хищника на популяцию жертвы осуществляется не с одинаковой интенсивностью в течение года. Обычно интенсивная гибель от хищников имеет место в течение сравнительно короткого промежутка времени, когда период активного питания хищника совпадает с таким состоянием жертвы, когда она оказывается относительно легко доступной для хищника. Это было выше показано на примере корюшки. У сома Silurus glanis L. дельты Волги вобла Rutilus rutilus caspicus Jak. играет важную роль в пище весной, с середины апреля до середины мая, когда сом съедает 68% своего годового рациона; летом в июне и июле основную пищу сома составляет молодь сазана Cyprinus carpio L., скатывающаяся из полоев в авандельту, а осенью - опять вобла, подходящая из моря в низовье Волги на зимовку. Таким образом, вобла в пище сома имеет значение лишь около двух месяцев - во время нерестового хода, нереста и во время миграции осенью на зимовку; в другое время сом в дельте Волги воблой практически не питается.
Иная картина наблюдается у жереха Aspius aspius (L.): он интенсивно поедает молодь воблы летом, когда она скатывается из нерестовых водоемов, главным образом в поверхностных слоях стрежневой части реки и недоступна для сома, но хорошо доступна для жереха. За летние месяцы (июнь-июль) жерех съедает 45% своего годового рациона, причем 83,3% (по числу штук) от всей пищи составляет молодь воблы. В остальное время года жерех воблой почти не питается (Фортунатова, 1962).
Щука, как и сом, поедает главным образом идущую на нерест воблу в нижней зоне дельты, где держатся более крупные щуки. Скатывающаяся молодь воблы для щуки, как и для сома, оказывается малодоступной (Попова, 1961, 1965).
Очень ограниченное время треска питается мойвой. Интенсивное питание трески мойвой продолжается обычно около месяца.
В Амуре малой корюшкой хищники обычно питаются интенсивно в два приема: весной, во время ее нереста, и осенью, во время ее миграции вверх по течению в прибрежной зоне (Вронский, 1960).
Очень сильно меняются условия воздействия хищников на жертвы в разные по гидрологическому режиму годы. В речных водоемах в многоводные годы обычно доступность жертв для хищников сильно снижается, а в годы с низким паводком увеличивается.
Определенное влияние оказывают хищники и на структуру популяции своей жертвы. В зависимости от того, на какую часть популяции воздействует хищник, он и вызывает соответствующую перестройку структуры популяции жертвы. Можно с уверенностью сказать, что большинство хищников осуществляет селективное изъятие особей из популяции. Лишь в некоторых случаях это изъятие не носит селективного характера, и хищник изымает добычу в том же размерном соотношении, в каком она содержится в популяции. Так, например, белуха Delphinapterus leucas, различные тюлени, калуга Huso dauricus (Georgi) и некоторые другие хищники выедают из стада ходовой кеты рыб без отбора определенных размеров. То же наблюдается, видимо, и в отношении скатывающейся молоди дальневосточных лососей - кеты и горбуши. Вероятно, неселективно питание трески нерестовой мойвой. В большинстве же случаев имеет место отбор хищником рыб определенного размера, возраста, а иногда и пола.
Причины избирательного питания хищников в отношении жертв разнообразны. Самая обычная причина - это соответствие относительных размеров и строения хищника размерам и строению, в частности наличию определенных защитных приспособлений, жертвы (шипов, колючек). Существенное значение имеет разная доступность разных полов. Так, например, у бычков, у колюшек во время охраны гнезда в большем количестве выедаются хищниками обычно самцы. Это отмечено, например, у Gobius paganellus (L.), что компенсируется большим процентом самцов у этого вида в потомстве (Miller, 1961). Меньшее выедание крупных рыб в нагульный период по сравнению с выеданием молоди часто может быть связано с их большей осторожностью (Милановский и Рекубратский, 1960). В целом же большинство рыб-хищников питается неполовозрелой частью стада жертвы. Половозрелая часть стада, особенно у крупных рыб, выедается хищниками сравнительно в незначительном количестве. В этом воздействие хищников отличается от воздействия вылова, который, как правило, изымает из популяции главным образом половозрелых особей. Так, из стада воблы хищники (судак, сом, щука) изымают рыб главным образом от 6. до 18 см длины, а промысел берет рыб длиной от 12 до 23-25 см (рис. 56).

Рис. 56. Размерный состав изымаемой из водоема воблы (по Фортунатовой, 1961): 1 - в уловах; 2 - в питании судака; 3 - в питании щуки; 4 - в питании сома; 5 - в питании сома, щуки и судака (суммарно)
Если к этому добавить выедание мальков воблы молодью хищных рыб, то разница получится еще более значительной (Фортунатова, 1961).
Таким образом, воздействие хищников на структуру популяции жертвы обычно сказывается путем выедания молоди, т. е. сокращения величины пополнения, что вызывает повышение среднего возраста половозрелой части популяции. Какую часть всего стада рыбы выедают хищники и какую относительную величину смертности популяция может компенсировать воспроизводством, мы знаем еще очень плохо. По-видимому, эта величина порядка 50-60% от нерестового стада у рыб с коротким жизненным циклом и 20-40% у рыб с длинным жизненным циклом и поздней половой зрелостью.
Количественных данных о том, какую часть популяции съели хищники, в литературе очень мало. Это затрудняется тем, что не удается определить общую численность ни популяции жертвы, ни хищника, ею питающегося. Однако в некоторых случаях попытки подобного рода делались. Так, Кроссмен (Crossman, 1959) определил, что радужная форель Salmo gairdneri Rich, выедает в оз. Пауль (Paul Lake) от 0,15 до 5% популяции Richardsonius balteatus (Rich.).
Иногда удается примерно определить соотношение естественной и промысловой смертности в отношении некоторых видов; так, К. Р. Фортунатова (1961) показала, что хищники съедают лишь немного меньше воблы, чем ее вылавливается промыслом (в 1953 г., например, было выловлено 580 тыс. ц воблы, а хищники съели 447 тыс. ц). Риккер (Ricker, 1952) выделяет три типа возможных количественных отношений хищника и жертвы:
1) когда хищник поедает определенное количество жертв, а остальная часть избегает поимки;
2) когда хищник выедает определенную часть популяции жертвы;
3) когда хищники поедают всех наличных особей жертвы, за исключением тех, которые могут избежать, поимки, спрятавшись в такие места, где хищник не может их добыть, или когда численность жертвы достигает столь малой величины, что хищник должен будет перейти в другое место.
В качестве примера первого случая, когда численность жертвы не ограничивает потребности хищника, Риккер приводит питание хищников нерестовыми скоплениями сельди или скатывающейся молодью лососей. В этом случае количество съеденных рыб определяется продолжительностью контакта с хищниками.
В качестве примера второго типа Риккер приводит поедание рядом хищников в оз. Култус молоди нерки, которой эти хищники питаются в течение всего года: здесь интенсивность выедания зависит как от численности жертвы, так и от численности хищника.
Наконец, третий случай, - когда интенсивность выедания определяется наличием убежищ и не зависит (естественно, в определенных пределах) от численности жертвы и численности хищника. Примером может служить выедание молоди атлантического лосося рыбоядными птицами в нерестовых реках. Как показано Элсоном (Elson, 1950,1962), вне зависимости от исходной численности популяции жертвы ее может выжить только такое количество, которое обеспечено убежищами, где жертва оказывается недоступной хищнику. Таким образом, количественное воздействие хищника на жертву может быть трояким: 1) когда съедаемое количество определяется продолжительностью контакта жертвы и хищника и численностью и активностью хищника; 2) когда количество съеденных жертв зависит как от численности жертвы, так и хищника и мало связано с временем контакта; 3) количество съеденных жертв определяется наличием необходимых убежищ, т. е. степенью доступности для хищника. Хотя эта классификация в известной степени формальна, но она удобна при разработке системы мер биотической мелиорации.
Воздействие хищника на жертву, его характер и интенсивность, как говорилось, специфичны для каждого этапа развития так же, как специфичны и формы защиты. У личинок китайского окуня основные органы защиты - это шипы на жаберной крышке, а у мальков - колючие лучи плавников в сочетании с высотой тела (Захарова, 1950). У мальков летучих рыб - это уплывание от преследователя и рассредоточивание, а у взрослых выскакивание из воды.
Воздействие большинства хищников обычно продолжается короткий промежуток времени как в течение года, так и суток, и знание этих моментов необходимо для правильной регуляции воздействия хищников на стадо промысловой рыбы.
|
|
|
