 |


Хозяйственное значение пресмыкающихся
|
|
|
|
Содержание
Характеристика класса "костные рыбы", систематика внутри класса. Морфология костных рыб
Подкласс "ящерохвостные архиоптерии"
Хозяйственное значение пресмыкающихся
Литература
Характеристика класса "костные рыбы", систематика внутри класса. Морфология костных рыб
Как и у хрящевых рыб, кожа костных рыб состоит из многослойного эпителия и подстилающего волокнистого соединительно-тканного кориума.
Многочисленные одноклеточные железы эпидермиса выделяют слизистый секрет, покрывающий тонким слоем тело рыбы. Эта слизистая пленка способствует уменьшению трения при плавании и, благодаря своим бактерицидным свойствам, препятствует проникновению в кожу бактерий. В одноклеточных железах кожи рыб, преимущественно мирных, содержится секрет, выделяющийся только при поранении кожи и служащий предупреждением об опасности - так называемое "вещество страха". При определенном состоянии рыб выделяемые кожными железами вещества - феромоны - стимулируют брачное поведение.
Клетки нижних слоев эпидермиса и клетки кориума содержат пигменты. Окраска большинства видов рыб криптична: она делает рыбу малозаметной на фоне места обитания. Придонные рыбы и рыбы травянистых и коралловых зарослей часто имеют яркую пятнистую или полосатую так называемую "расчленяющую" расцветку, маскирующую контур рыбы. Характерная для донных рыб дорзо-вентральная уплощенность тела уменьшает размер отбрасываемой тени и также облегчает маскировку. Некоторые рыбы способны произвольно менять окраску в зависимости от цвета субстрата.
Особенно отчетливо эта способность выражена у камбаловых, некоторых бычков, скорпен и других рыб. Смена окраски происходит в течение нескольких минут. Ослепленные рыбы приобретают темную окраску и в дальнейшем ее не меняют. Смена окраски обеспечивается изменением формы хроматофоров (пигментных клеток) под влиянием импульсов, поступающих из продолговатого мозга через симпатическую нервную систему (центр "посветления") и из промежуточного мозга (центр "потемнения") через парасимпатическую систему. Такая двойная антагонистическая иннервация обеспечивает быстрые изменения окраски.
|
|
|
У пелагических рыб темная окраска спины делает их малозаметными на фоне дна при рассматривании сверху, а блестящая серебристая окраска боков и брюшка делает рыбу незаметной на фоне светлого неба при рассматривании снизу. У многих пелагических рыб (сельдевые, некоторые карповые и другие) брюшко сжато в виде острого киля, ликвидирующего демаскирующую тень на брюхе при освещении рыбы сверху и усиливающего маскирующее значение серебристой окраски нижней части тела (рис.1)
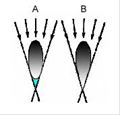
Рис.1. Схема образования демаскирующей тени (A, область голубого цвета) и исчезновения тени при брюшном киле (B) рыб.
У некоторых видов (колюшки, лососевые и др.) в сезон размножения развивается яркая брачная окраска. Ее появление обусловлено действием гормонов и служит важным условием развития полового поведения и синхронизации созревания половых продуктов у самцов и самок.
У большинства видов костных рыб в коже образуются защитные костные образования - чешуи; у части видов они редуцируются. Многие ископаемые кистеперые (и ныне живущая латимерия) и двоякодышащие рыбы имели космоидную чешую в виде костной пластинки, снаружи покрытой слоем более плотного костного вещества - космина (видоизменение дентина). По происхождению космоидная чешуя представляет собой комплекс слившихся и сильно измененных плакоидных чешуй.
|
|
|
костная рыба пресмыкающийся морфология
Космоидная чешуя строится из глубоко расположенного слоя изопедина. Средний слой представлен губчатой костью, а поверхность - из космина, в виде приплюснутых дентина зубчиков, между которыми располагаются специфические колбовидные полости, открывающиеся наружу порами, электрорецепторы.
В эволюции из космоидной возникла ганоидная чешуя. Последняя состоит из костной пластинки, сверху покрытой дентиноподобным веществом - ганоином. Обычно ганоидные чешуи имеют ромбическую форму и, соединяясь друг с другом специальными сочленениями, образуют панцирь, одевающий все тело рыбы. Такая чешуя была свойственна ископаемым палеонискам, а из ныне живущих рыб ее сохранили многоперообразные (имеют космо-ганоидную чешую) и панцирникообразные (чешуя ганоидная). Как и космоидная, ганоидная чешуя закладывается в кориуме. При детальном изучении под микроскопом выяснилось, что ганоидная чешуя сверху покрыта многослойным ганоином, глубже находится космин, который подстилается изопедином. Ганоидная чешуя с космином называется палеонискоидной, а без космина - лепидостеоидная.
Костные чешуи, свойственные остальным современным костным рыбам, филогенетически представляют видоизменение ганоидной чешуи, у которой исчез поверхностный слой ганоина (в виде очень тонкого слоя ганоин сохраняется на костной чешуе амии). Костные чешуи образуются в поверхностных слоях кориума; обычно они черепицеобразно накладываются друг на друга. У большинства рыб чешуи циклоидные - с гладким наружным краем. У окунеобразных и некоторых других рыб по заднему (наружному) краю чешуйки развиты зубчики - это ктеноидная чешуя. Чешуи растут в течение всей жизни рыбы. Верхняя поверхность чешуи часто имеет сложный рельеф (ребристость, исчерченность), увеличивающий ее прочность и направляющий (организующий) обтекающие тело рыбы потоки воды. На боковых сторонах хвостового стебля таких быстрых пловцов, как многие скумбриевые, некоторые сельдевые и др., расположены группы чешуи с очень сложным поверхностным рельефом. У многих рыб в нижних слоях чешуи лежит прослойка кристалликов извести и серебристого пигмента - гуанина, усиливающих блестящую окраску рыбы. При потере чешуи (механические повреждения и т.п.) довольно быстро (через 20-50 дней) происходит их полная регенерация.
|
|
|
В соответствии с неравномерным ростом рыбы в течение года на чешуйке образуются концентрические кольца: широкие соответствуют периоду интенсивного роста, узкие - образуются в сезон его замедления. Это позволяет использовать чешую для определения возраста рыбы (с той же целью используется слоистость и некоторых костей - клейтрума, жаберной крышки и др.). Длина чешуи и длина тела рыбы с возрастом изменяются пропорционально друг другу. Основываясь на этом, были разработаны методики, позволяющие рассчитывать скорость роста данной особи за предыдущие годы.
Помимо чешуи в более глубоких слоях кориума образуются покровные кости черепа и пояса грудных плавников (см. ниже). Кожа костных рыб, как и хрящевых, проницаема для воды и некоторых растворенных в ней веществ.
Скелет и мышечная система. У костных рыб хрящ в скелете в той или иной степени замещается костной тканью: образуются основные или замещающие кости. Кроме этого, в коже возникают покровные кости, погружающиеся потом под кожу и входящие в состав внутреннего скелета. Скелет костных рыб подразделяется на осевой скелет, череп (мозговой и висцеральный), скелет непарных плавников, скелет парных плавников и их поясов.
У кистеперых, двоякодышащих и осетрообразных рыб функцию опоры в осевом скелете выполняет хорда, окруженная плотной соединительнотканной оболочкой. Хорошо развитые, иногда частично окостеневающие верхние дуги образуют канал, в котором лежит спинной мозг. К слабо развитым нижним дугам прикрепляются ребра. У части предков этих групп встречались более или менее развитые тела позвонков. У многоперообразных и всех костистых рыб хорошо развиты костные позвонки амфицельного типа (двояковогнутые). Сильно редуцированная хорда имеет четкообразное строение: она расширена в пространстве между телами позвонков и в сильно суженном виде проходит через канал в центре тела позвонка.
|
|
|
Позвонки туловищного отдела несут костные верхние дуги, заканчивающиеся длинными верхними остистыми отростками; к поперечным отросткам тел позвонков причленяются длинные и тонкие костные ребра, хорошо развитые у большинства костных рыб. Позвонки хвостового отдела несут верхние дуги с остистыми отростками, а поперечные отростки смещаются книзу и, сливаясь попарно, образуют нижние дуги, замкнутые нижними отростками
Промежутки между дугами затянуты плотной соединительнотканной пленкой. В верхнем канале лежит спинной мозг; нижние дуги хвостовых позвонков образуют гемальный канал, в котором идут хвостовые артерия и вена; защищенные от сдавливания мощными мышцами этого отдела. Позвонки соединяются друг с другом при помощи суставных отростков, расположенных у основания верхних дуг. Такие сочленения обеспечивают прочность осевого скелета при сохранении его подвижности. Позвоночный столб может изгибаться преимущественно в горизонтальной плоскости. У большинства костистых рыб в толще мышц лежат тонкие мускульные косточки, создающие дополнительную опору для мускульных волокон.
Череп костных рыб, как и всех позвоночных, подразделяется на мозговой (осевой) и висцеральный отделы.
У осетрообразных мозговой череп остается хрящевым; лишь у старых рыб в нем появляются небольшие окостенения. Снаружи хрящевой череп покрыт сплошным панцирем из большого числа покровных костей. Но для большинства костных рыб характерно замещение хряща мозгового черепа основными костями, непосредственно прилегающими друг к другу или соединенными остатками хряща, и относительно небольшое число покровных костей - остатков первичного панциря. Окостенение мозгового черепа сильно развито у ганоидных (кроме осетрообразных) и костистых рыб. У кистеперых и двоякодышащих рыб в черепе сохраняется большое количество хряща и развиваются лишь немногие основные кости; первичный панцирь из покровных костей у них хорошо развит.
У костистых рыб в затылочной части образуются четыре окаймляющие большое затылочное отверстие кости - основная (basioccipitale), две боковые (occipitale laterale) и верхняя (supraoccipitale) затылочные. В боковой стенке черепа 5 ушных костей (ossa otici). В области глазницы возникают клиновидные кости: глазоклиновидная (orbitosphenoideum), основная (basisphenoideum) и боковая клиновидная (laterosphenoideum). В области обонятельного отдела формируется непарная средняя (mesethmoideum) и парные боковые обонятельные кости (ectoethmoideum). Все эти кости основные: развиваются путем окостенения участков хряща. Сверху череп прикрывают 3 пары покровных костей: носовые (nasale), очень большие лобные (frontale) и маленькие теменные (parietale). Дно мозгового черепа образовано двумя непарными покровными костями: большим парасфеноидом (parasphenoideum) и несущим зубы сошником (vomer). Благодаря сложному наружному рельефу костей границы между ними не всегда видны.
|
|
|
Висцеральный скелет черепа представляет собой систему хрящевых или в разной степени окостеневших дуг - челюстной, подъязычной и 5 жаберных; четыре покровные кости образуют жаберную крышку. Покровные кости укрепляют челюстную дугу, образуя вторичные челюсти. Для костных рыб характерна гиостилия: соединение челюстной дуги и вторичных челюстей с мозговым черепом через верхний элемент подъязычной дуги - подвесок или гиомандибуляре (hyomandibulare). Только у двоякодышащих верхняя челюсть срастается с дном мозгового черепа (аутостилия), а подвесок, утративший свою функцию, уменьшается в размерах.
У осетрообразных в висцеральном скелете сохраняется много хряща, а вторичные челюсти слабо развиты. У костистых рыб в этом отделе хрящ полностью замещен костями.
В результате окостенения первичной верхней челюсти - небно-квадратного хряща - с каждой стороны образуется несущая зубы небная кость (palatinum), а в задней его части - задняя крыловидная (metapterygoideum) и квадратная (quadratum) кости. Между ними располагаются покровные наружная и внутренняя крыловидные кости (ectopterygoideum и entopterygoideum). Первичная нижняя челюсть - меккелев хрящ, окостеневая, превращается в сочленовную кость (articulare), образующую с квадратной костью челюстной сустав. У костистых рыб хорошо развиты вторичные челюсти из покровных костей; они срастаются или соединяются связками с костями первичных челюстей. В верхней челюсти такими вторичными образованиями будут предчелюстная (praemaxillare) и верхнечелюстная (maxillare) кости; на них сидят зубы (у части видов отсутствуют на одной или обеих костях). Мощная покровная зубная кость (dentale) образует основную часть нижней челюсти. Функцию схватывания и удерживания добычи выполняют как первичные, так и вторичные челюсти.
Мощность и характер зубов, равно как относительные размеры челюстей и положение ротового отверстия, отражают пищевую специализацию каждого вида.
Подъязычная дуга образована основными костями. Ее верхний элемент - подвесок, представлен крупной костью hyomandibulare, широким верхним краем причленяющейся к слуховому отделу осевого черепа. Через отщепившуюся от нижнего конца подвеска дополнительную косточку (syniplecticum) гиомандибуляре причленяется к квадратной кости (гиостилия), а через короткую окостеневшую связку interhyale - к нижнему элементу подъязычной дуги - гиоиду, в котором образуется несколько окостенений, часто сливающихся в общую кость (hyoideum). Передние концы гиоидов правой и левой сторон соединены друг с другом непарной косточкой - копулой (copula), поддерживающей складку языка. К гиоиду прикрепляются тонкие изогнутые косточки - лучи жаберной перепонки, поддерживающие кожистый край жаберной крышки. Жаберная крышка у костных рыб образована покровными костями. К заднему краю подвеска и квадратной кости плотно прилегает мощная, резко изогнутая предкрышечная кость (praeoperculum), к которой присоединяются крышечная (operculum), межкрышечная (interoperculum) и подкрышечная (suboperculum) кости.
Жаберных дуг 5 пар. Первые четыре состоят из 4 окостеневших и соединенных друг с другом суставами, парных элементов; расположенные снизу пятые непарные элементы соединяют дуги друг с другом. Эти жаберные дуги несут жабры. Пятая (задняя) жаберная дуга состоит только из двух крупных парных элементов; на них у некоторых видов расположены глоточные зубы (форма и размеры глоточных зубов отражают пищевую специализацию).
Парные конечности и их пояса. Парные конечности костных рыб представлены грудными и брюшными плавниками. У немногих видов брюшные плавники редуцируются. В подклассе лопастеперых рыб внутренний скелет парных грудных и брюшных плавников имеет центральную расчлененную ось, к которой причленяются радиалии (они могут в разной степени редуцироваться). У двоякодышащих рыб радиалии прикрепляются по бокам члеников хорошо развитой центральной оси попарно (бисериальный тип плавника). Таков скелет плавников неоцератода и многих ископаемых форм. У остальных современных двоякодышащих рыб радиалии редуцируются и нитевидный плавник сохраняет лишь расчлененную центральную ось.
У кистеперых рыб радиалии прикрепляются к менее четко выраженной членистой центральной оси только с одной стороны (унисериальный тип). Внутренний скелет плавников у лопастеперых остается хрящевым, но у ископаемых форм в нем развивались окостенения. Краевая часть плавника поддерживается наружным скелетом - лепидотрихиями - костными лучами кожного происхождения. В подклассе лучеперых внутренний скелет парных плавников более простой.
У многоперообразных в основании грудного плавника лежат три частично окостеневающих базальных элемента; к ним прилегает ряд мелких радиалии, к которым причленяются основания костных кожных лучей - лепидотрихий. У остальных лучеперых рыб базальные элементы редуцируются. В грудных плавниках небольшое число окостеневших радиалии прикрепляется к поясу; к радиалиям, а частично и непосредственно к поясу, причленяются липидотрихии. В брюшных плавниках внутренний скелет редуцируется, и лепидотрихий причленяются непосредственно к тазовому поясу (несколько радиалий сохраняются у немногих видов).
Первичный пояс грудных плавников у осетрообразных имеет вид. Двух хрящевых полудуг, снаружи покрытых несколькими кожными костями, образующими вторичный пояс. У двоякодышащих рыб кости вторичного пояса погружаются под кожу и тесно прилегают к первичному хрящевому поясу, значительно его укрепляя. У остальных групп костных рыб первичный пояс окостеневает, образуя лопатку (scapula) и коракоид (coracoideum). Обычно они относительно невелики и фактически выполняют функцию редуцировавшихся базальных элементов внутреннего скелета плавника. Функцию пояса выполняют только вторичные кости. Из них особенно велик клейтрум (сleithrum), который через добавочные косточки supracleithrum - posttemporale причленяется к затылочному отделу черепа.
Клейтрумы правой и левой стороны на брюшной стороне тела соединяются друг с другом. Эти преобразования обеспечивают более прочную, чем у хрящевых рыб, фиксацию пояса грудных плавников.
Пояс брюшных плавников у костных рыб развит слабо. Он представляет собой либо хрящевую (двоякодышащие, осетрообразные| либо костную пластинку, лежащую в толще мускулатуры. У части костистых рыб брюшные плавники вместе со своим поясом перемещаются вперед и располагаются под грудными (иногда немного впереди). В этом случае пояс брюшных плавников также лежит в толще брюшной стенки, но при помощи связок соединяется с поясом грудных плавников.
Внутренний скелет непарных плавников образован палочковидными хрящевыми или костными радиальными лучами - птеригиофорами (pterygiophora), погруженными в толщу мышц. Их заостренные нижние концы общей связкой соединяются с остистыми отростками позвонков. К лежащим на уровне поверхности тела расширенным верхним концам птеригиофоров подвижно причленяются кожные костные лучи - лепидотрихии (lepidotrichia), поддерживающие лопасть плавника.
У многих видов часть или все лепидотрихии непарных плавников становятся мощными и острыми; такие плавники выполняют не только гидродинамическую, но и защитную функцию (оку необразные и представители других отрядов). Иногда часть лучей обособляется и превращается в отдельно стоящие мощные подвижные колючки (колюшки, некоторые сомы и др.). У немногих видов у основания колючек или части колючих лучей плавников расположены ядовитые железы: при надавливании на колючку выделяется капелька яда, по колючке стекающая в ранку (морские окуни и ерши, морской дракончик и др.). У видов, живущих в горных ручьях и в прибрежных участках морей с сильными приливно-отливными течениями, брюшные плавники, видоизменяясь, превращаются в присоску, которой рыба прикрепляется к субстрату (бычки-Gobiidae, подкаменщики-Cottidae, пинагоры - Cyclopteridae, некоторые сомы и др.). У рыбы прилипалы первый спинной плавник сместился на голову и превратился в сложно устроенную мощную присоску. У морских петухов из отряда скорпенообразные три луча каждого грудного плавника служат рыбе опорой при ползании по дну.
Хвостовой плавник у личинок рыб имеет симметричное строение (хорда проходит посредине плавника) и называется протоцеркальным (рис.3). У осетрообразных, как и у хрящевых рыб, конец хвоста изгибается вверх, а брюшная лопасть разрастается; образуется неравнополостный, или гетероцеркальный, плавник.
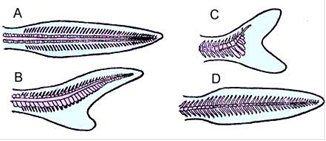
Рис.3. Форма хвостового плавника. A - протоцеркальная; B - гетероцеркальная (осетрообразные); C - гомоцеркальная (большинство костистых рыб); D - дифицеркальная
У большинства костистых рыб брюшная лопасть разрастается еще сильнее, плавник становится внешне симметричным, но конец позвоночника заходит в верхнюю лопасть; плавник такого типа называется гомоцеркальным. У части двоякодышащих, кистеперых и немногих костистых рыб при росте осевой скелет вновь выпрямляется, а верхняя и нижняя лопасти хвостового плавника уравниваются в размерах; такой вторично симметричный хвостовой плавник называют дифицеркальным. Лопасть хвостового плавника поддерживают костные лучи-лепидотрихии, а функцию его внутреннего скелета выполняют расширенные остистые отростки концевых позвонков; у костистых рыб нижние остистые отростки разрастаются в широкие пластинки (гипуралии).
Как и у хрящевых рыб, поперечнополосатая соматическая мускулатура костных рыб отчетливо сегментарна и состоит из мышечных сегментов, имеющих сложную форму. Часть из них частично перестроилась, образовав группы мышц: глазные, наджаберные и поджаберные мышцы, мышцы парных плавников.
Висцеральная мускулатура, окружающая пищеварительную трубку представлена слоями гладких мышц, но у некоторых костистых рыб среди гладких мышц встречаются пучки поперечнополосатых волокон (в стенках желудка вьюна, в кишечной стенке линя и др.). В области челюстных и жаберных дуг гладкие мышечные волокна замещаются поперечнополосатыми волокнами, формирующими сложно дифференцированную мускулатуру челюстных и жаберных дуг. В отличие от хрящевых рыб содержание мочевины в мышцах костных рыб ничтожно и обычно не превышает 0.0005 - 0.02%.
Подкласс "ящерохвостные архиоптерии"
Птиц принято выделять в особый класс Aves, но во многом они отошли от общего ствола рептилий не больше, чем их предки - некоторые из архозавров. Среди архозавров была одна группа летающих форм - птерозавры. Однако предками птиц были не птерозавры, а летающие архозавры другого типа, которые вместо перепонок использовали для полета измененные передние конечности - крылья, покрытые перьями (диагностический признак класса птиц).
В некоторых отношениях, особенно по адаптациям, связанным с двуногостью, птицы сходны со своими родичами среди динозавров, но почти каждый специфический признак птиц представляет собой адаптацию к полету.
Высокая и постоянная температура тела и более совершенная система кровообращения связаны с необходимостью поддерживать высокую интенсивность обмена, способную обеспечить длительный полет. Уменьшение веса тела различными способами (главным образом за счет развития воздушных мешков и пневматизации костей) также связано с полетом, равно как и модификации в головном мозге и в органах чувств.
В процессе эволюции птицы давным-давно открыли законы аэродинамики, которые человеку пришлось постигать ценой долгих исследований и горького опыта; птицы - великие "знатоки" во всем, что касается строения крыла и использования ветра и воздушных течений для того, чтобы выиграть в расстоянии и высоте. В общем у птиц с активным машущим полетом крылья относительно короткие и широкие; у птиц с парящим или планирующим полетом поверхность крыльев обычно больше, а длина в большинстве случаев очень велика.
Летающие птицы (в отличие от бегающих по земле, подобно страусам) редко бывают крупными. Для того чтобы птица могла летать, площадь крыльев должна возрастать в общем пропорционально массе тела, а при слишком крупных размерах потребовался бы такой большой размах крыльев, что ими стало бы трудно манипулировать.
Все птицы, за исключением примитивных мезозойских форм, утратили зубы, и функцию сбора пищи выполняет клюв. Строение клюва отличается большим разнообразием - от массивного клюва попугая до эффективного долбящего клюва дятла. По-видимому, примитивные птицы питались какой-то сравнительно мягкой пищей, поэтому потребность в зубах у них отпала. Однако многие современные птицы питаются семенами. Для такой пищи млекопитающим нужны сильно развитые перетирающие зубы; у птиц же функцию перетирания пещи выполняет мускульный желудок, выстланный изнутри роговой кутикулой и содержащий мелкие камушки. Мускульный желудок имеется также и у крокодилов; вполне возможно, что такой желудок - признак, общий для всех архозавров.
По счастливой случайности в палеонтологической летописи сохранились пять скелетов предка птиц Archaeopteryx - единственного представителя подкласса Archaeornithes из верхнеюрских отложений. У этой птицы еще имелись зубы, а также вооруженные когтями пальцы на крыльях и длинный рептильный хвост; в строении скелета археоптерикса так равномерно представлены признаки архозавров и современных птиц, что его положение в системе оставалось бы неясным, если бы вместе со скелетом не сохранились отпечатки перьев. Сравнение археоптерикса с различными вымершими архозаврами позволяет считать, что он, как и птицы вообще, происходит от мелких тероподных динозавров, однако, по этому вопросу существуют разногласия, и некоторые палеонтологи считают предками птиц текодонтов, птицетазовых динозавров и даже древних крокодилов.
Хозяйственное значение пресмыкающихся
Пресмыкающиеся по сравнению с земноводными представляют собой следующий этап приспособления позвоночных животных к жизни на суше. Это первые настоящие наземные позвоночные, характеризующиеся тем, что размножаются на суше яйцами, дышат только легкими, механизм дыхания у них всасывательного типа (при помощи изменения объема грудной клетки), хорошо развиты проводящие дыхательные пути, кожа покрыта роговыми чешуйками или щитками, кожных желез почти нет, в желудочке сердца имеется неполная или полная перегородка, вместо общего артериального ствола от сердца отходят три самостоятельных сосуда, почки тазовые (метанефрос). У пресмыкающихся увеличивается подвижность, что сопровождается прогрессивным развитием скелета и мускулатуры: меняется положение различных отделов конечностей по отношению друг к другу и к телу, укрепляются пояса конечностей, позвоночник расчленяется на шейный, грудной, поясничный, крестцовый и хвостовой отделы, увеличивается подвижность головы. Череп пресмыкающихся, как и у птиц, в отличие от других позвоночных животных, соединяется с позвоночником одним (непарным) мыщелком. В скелете свободных конечностей характерны межзапястные (интеркарпальные) и межпредплюсневые (интертарзальные) сочленения. В поясе передних конечностей у них есть своеобразная покровная кость-надгрудинник. К примитивным особенностям пресмыкающихся как обитателей суши следует отнести наличие двух дуг аорты, смешанную кровь в артериях туловищного отдела, низкий уровень обмена веществ и непостоянную температуру тела. Современные пресмыкающиеся представляют собой лишь разрозненные остатки богатого и разнообразного мира рептилий, населявшего Землю в мезозойскую эру.
В пустынях и степях, где численность пресмыкающихся высока, они играют заметную роль в биоценозах. Большинство ящериц и змей, уничтожая значительное количество вредных насекомых, моллюсков и грызунов, приносит пользу сельскому хозяйству. Многие ящерицы в пустынях служат кормом для ряда важных промысловых зверей, например лисицы и хорька. Некоторые виды приносят местами заметный вред. Среднеазиатская черепаха в ряде районов оказывается вредителем на бахчах и особенно в питомниках фисташки; вредная деятельность этой черепахи усугубляется прокапыванием ею насыпей и стен оросительной системы. На рыборазводных станциях серьезный вред приносят водяные ужи, уничтожающие большое количество мальков промысловых рыб: карпа, лосося, осетра. В районах рыборазведения водяные ужи подлежат полному уничтожению, что лучше всего делать ранней весной на местах их зимовок. Вероятна отрицательная роль ряда пресмыкающихся, особенно сухопутных черепах, некоторых ящериц и змей, в распространении ряда заболеваний человека и домашних животных, поскольку на них живут нимфы и взрослые иксодовые клещи. В некоторых странах серьезный вред человеку приносят ядовитые змеи.
Количество жертв от ядовитых змей обычно сильно преувеличено, но всё же в жарких странах, особенно в Юго-Восточной Азии и Южной Америке, ежегодно от укусов змей умирают тысячи людей. В Западной Европе далеко не ежегодно регистрируются единичные случаи смерти людей от укусов ядовитых змей. В Средней Азии, где наблюдается наибольшая численность ядовитых змей, ежегодно от их укусов умирает до 12 человек и смертельные случаи составляют около 6% укушенных (главным образом дети, укушенные в лицо). Прежде, до применения сывороток, укусы наиболее ядовитых змей - кобры, гюрзы, эфы - давали от 15 до 30% смертельных случаев, гадюки - 5-6%. Сейчас смертельные исходы бывают в 1,5-2% случаев укусов. В зависимости от характера действия яда различают две группы ядовитых змей. Человек, укушенный змеей I группы, к которой относятся гюрза, эфа, гадюка, ощущает сильную боль, окружающая ранку ткань краснеет и опухает. Развивающийся отек распространяется от места укуса на всю конечность. Появляется сонливость, выступает холодный пот. В ряде случаев бывает лихорадочное состояние, тошнота, бред. Вследствие подкожных точечных кровоизлияний на теле часто появляются фиолетово-красные пятна. В тяжелых случаях наступает некроз тканей, образуются нарывы и язвы. Может наступить смерть. Человек, укушенный змеей II группы, яд которой действует через нервную систему (к ним относится кобра), обычно не чувствует боли в месте укуса. Однако вскоре после укуса развивается слабость в конечностях - начинается паралич. Распространяясь, паралич захватывает все органы, затрудняет дыхание, речь, опускаются веки, парализуются губы, язык. Лицо синеет. В тяжелых случаях наступает смерть. Большое значение для лечения укушенного имеет своевременное оказание помощи. С укушенного места следует снять остатки яда крепким раствором марганцовокислого калия, давать больному крепкий чай, кофе и обеспечить полный покой и тепло. Больного необходимо как можно скорее доставить в больницу, где ему будет введена специальная сыворотка. Сыворотка хранится в ампулах или в сухом виде с указанием дозировки.
Обычно вводят под кожу 10 см3, но в тяжелых случаях прибегают к венозным инъекциям большими дозами. Увеличивать ранку надрезами для усиления кровотечения бесполезно. Только вред могут принести прижигание, прикладывание всевозможных "змеиных" камней, земли и т.д. Наложение жгута - опасное и практически также бесполезное средство, так как оно не задерживает всасывания яда. В большинстве случаев тяжелые последствия после укуса ядовитых змей бывают вызваны не действием яда, а прижиганиями, кровопусканиями, перетягиванием конечностей жгутом выше места укуса и т.д. Змеиный яд применяется для приготовления лечебных сывороток против укусов змей. Сыворотка приготавливается из крови лошади, в организм которой вводят возрастающие дозы яда в течение 16 месяцев. В результате кровь приобретает такой иммунитет, что животное может переносить впрыскивание 80-кратной смертельной дозы. Изготовляют моновалентные сыворотки, специфичные по отношению к яду одного вида змей, и поливалентные, действующие против яда нескольких видов змей. Из яда гадюк приготавливают также лечебные препараты, например випратокс, випраксин, випразид и другие, применяемые при лечении радикулита, ишиаса, ревматизма. Из яда гюрзы и щитомордника изготовляют кровоостанавливающие средства (лебетоке), из яда гремучей змеи - препараты, излечивающие эпилепсию, яд кобры употребляют для изготовления препаратов, применяемых при лечении бронхиальной астмы и ряда других заболеваний. Яд змей широко применяется в биохимии. Для получения яда организуют питомники змей. Первый такой питомник - серпентарий - был создан в Бразилии около города Сан - Пауло (Бутантан); он и сейчас самый крупный в мире. Подобные питомники имеются в Индии, Индонезии, Южной Африке и других странах.
В питомниках яд у змей берут постоянно, не убивая их. Пресмыкающиеся служат источником сырья для промышленности. Издавна кожа крокодила, крупных змеи и ящериц идет на изготовление чемоданов, портфелей, обуви и т.д. Особенно ценится кожа крокодила. В начале этого века крокодилов заготовляли по 500 тыс. экземпляров в год. Местами численность их настолько сократилась, что они были взяты под охрану и сделаны попытки искусственного разведения их. Так, фермы Флориды содержали до 12 тыс. крокодилов, поставляя на рынки до 3000 экземпляров в год. Среди наших видов как сырье для кожевенной промышленности имеют значение вараны и крупные полозы. Используется панцирь некоторых морских черепах, особенно биссы, роговые щитки которых в горячей воде и под давлением спрессовываются в однородную массу. Из этой высококачественной по пластическим свойствам красивой массы изготовляют гребни, оправы для очков и т.д. Некоторые рептилии употребляются человеком в пищу, наиболее часто мясо и яйца черепах. Существует регулярный промысел мягкокожистых черепах и некоторых видов морских черепах, особенно зеленых. В Северной Америке промышляют террапинов, которых стали разводить и на фермах. Среди наших видов вполне съедобно мясо средиземноморской, среднеазиатской и дальневосточной мягкокожистой черепах. Можно употреблять в пищу желтки яиц всех наших черепах. Мясо черепах идет также на откорм пушных зверей.
Литература
1. А. Ромер, Т. Парсонс "Анатомия позвоночных", издательство "Мир", 2003
2. Дзержинский Ф.Я. "Сравнительная анатомия позвоночных животных" Издательство: "Аспект Пресс" 2009
|
|
|
