 |


Пойкилотермность и гомойтермность
|
|
|
|
У животных реакции на разный тепловой режим жизнеобеспечения не менее разнообразны, чем у растений. И все они направлены на регулирование уровня теплопередачи. В отличие от растений для животных характерны два типа теплообмена: пойкилотермность (poikilos – разнообразный) и гомойтермность (homois – одинаковый).
К пойкилотермным (эктотермным, устаревшее – холоднокровным) относятся все беспозвоночные, рыбы, рептилии и амфибии. Они лишены способности поддерживать постоянную температуру тела. Для пойкилотермных организмов типична низкая интенсивность обмена веществ и почти полное отсутствие механизмов теплорегуляции. В тропических странах они встречаются чаще, чем в других.
Терморегуляция осуществляется за счет особой структуры и цвета покровов, специфики поведения – отыскивают наиболее подходящие местообитания (змеи выползают на скальные выходы, ящерицы – на стволы деревьев с солнечной стороны, лягушки – на теплые камни, листья), усилением мускульной работы (в полете – на 15-20°С температуры выше окружающей среды; у шмелей на Кавказе в горах – до 38-40°С при 4-8°С воздуха); за счет общественной жизни (муравейники, термитники, ульи); разным содержанием влаги в теле и разной интенсивностью испарения влаги с поверхности тела (эти наиболее безразличны к любым изменениям температуры воздуха); и др. Устойчивость к низким температурам обеспечивается накоплением жиров, гликогена, некоторых солей. Неблагоприятные условия пойкилотермные животные переживают в неактивном состоянии – анабиозе.
Гомойтермные (эндотермные, теплокровные) – животные с высоким уровнем обменных процессов – птицы и млекопитающие, обеспечивающими поддержание постоянной температуры тела даже при значительных колебаниях температуры внешней среды. Тепло выделяется при биохимических реакциях внутри организма. Чем ниже температура среды, тем больше потери тепла и тем интенсивнее идут обменные процессы, повышается продуцирование тепла, идущего на поддержание постоянной температуры тела. Аналогичная закономерность и при повышении температуры. Но эта закономерность прослеживается лишь до определенного предела. Ресурсы организма не беспредельны. При длительном перегреве или переохлаждении он погибает.
|
|
|
У гомойтермных животных различают химическую и физическую терморегуляции. Химическая проявляется в продуцировании тепла, физическая – в его распределении по телу и отдаче. У животных перед наступлением холодов возрастает в тканях печени содержание гликогена, в почках – аскорбиновой кислоты. Наблюдается накопление жиров под кожей и вблизи жизненно важных органов – сердца, спинного мозга. Жиры откладываются в особой бурой жировой ткани, и при клеточном дыхании вся энергия идет не на синтез АТФ, а рассеивается по телу в виде тепла.
Поддержание температуры за счет возрастания теплопродукции требует большого расхода энергии, поэтому животные при усилении химической терморегуляции либо нуждаются в большом количестве пищи, либо тратят много жировых запасов, накопленных ранее. Например, бурозубка крошечная имеет исключительно высокий уровень обмена. Чередуя очень короткие периоды сна и активности, она деятельна в любые часы суток, не впадает в спячку зимой и в день съедает корма в 4 раза больше собственной массы. Частота сердцебиения у бурозубок до 1000 ударов в минуту. Также и птицам, остающимся на зиму, нужно много корма; им страшны не столько морозы, сколько бескормица.
На основе физиологических процессов осуществляется терморегуляция в пределах тела: в конечностях вены и артерии подходят близко друг к другу («чудесная сеть») и артерии отдают тепло венам, возвращая его телу. В результате конечности остаются более холодными по сравнению с телом. В жару поддерживать температуру тела на постоянном уровне позволяет потоотделение, учащенное дыхание (собаки, птицы).
|
|
|
У экологически близких млекопитающих в холодных климатических зонах, согласно правилу Бергмана, закономерно увеличиваются размеры тела и вес внутренних органов, имеющих отношение к регулированию процессов обмена (сердце, почки, печень). Согласно правилу Аллена, в холодных зонах относительно размера тела сокращаются площади поверхностей выступающих органов (уши, носы, хвосты) по сравнению с млекопитающими более теплых зон. Правило Аллена наглядно демонстрируют размеры ушей у песца (Арктика), европейской лисы и африканской лисы-фенека.
Снижению теплопотерь способствуют опушение (как и у растений), оперение, шерстный покров, жировые отложения, темный окрас покрова (правило Глогера).
Промежуточное положение между пойкилотермными и гомойтермными организмами занимают гетеротермные (суслики, ежи, летучие мыши, медведи). В активном состоянии у этих животных поддерживается постоянная относительно высокая температура тела. В зимнее время они впадают в спячку или глубокий сон, и температура тела у них в это время мало отличается от внешней. Уровень обмена веществ снижается.
Температура и влажность являются ведущими климатическими факторами и тесно взаимосвязаны между собой. При неизменном количестве воды в воздухе относительная влажность увеличивается, когда температура падает. Если воздух охлаждается до температуры ниже точки водонасыщения (100%), происходит конденсация и выпадают осадки.
Влага в жизни организмов
Протекание всех биохимических процессов в клетках и нормальное функционирование организма в целом возможны только при достаточном обеспечении его водой — необходимым условием жизни.
Наиболее высоко содержание воды в периоды активной жизнедеятельности и в молодом возрасте.
Таблица 1 Содержание воды в различных организмах, % от массы тела
| Растения | Содержание воды | Животные | Содержание воды |
| Водоросли Морковь Разнотравье Листья деревьев Стволы деревьев Картофель (клубни) | 96-98 87-91 85-86 79-82 40-65 74-80 | Губки Моллюски Насекомые Ланцетник Земноводные Млекопитающие (мышечные ткани) | 80-92 46-92 до 93 68-83 |
Но и в состоянии покоя растения не теряют влагу полностью. В сухих лишайниках содержится до 5-7% воды, в зерновках злаков – 12-14%. Независимо от климата и почвенных условий в течение года всегда можно выделить такие периоды в развитии растений, за исключение растений влажных тропиков, когда они испытывают дефицит влаги. При остальных благоприятных условиях он сильно сказывается на росте и развитии растений, обусловливает их низкорослость и бесплодие.
|
|
|
В процессе эволюции у растений и животных выработался многочисленные сложные приспособления, позволяющие поддерживать водный баланс и обеспечивать экономное расходование воды. Растения пустынь и степей приспособились к острому дефициту влаги, болотные и влажно-тропические растения – к избытку, а лесным видам необходима высокая влажность воздуха и умеренная влажность почв. Как и в отношении остальных факторов, эти приспособления-адаптации группируются в анатомо-морфологические, физиологические и поведенческие.
Источниками влаги для растений служат запасы ее в почве и атмосфере (осадки, туманы, конденсаты), для наземных животных – вода в водоемах, водяные пары в атмосфере и сочная пища. При анализе влияния влаги на живые организмы важно учитывать сезонное распределение и температурный режим среды обитания. Разные комбинации содержания воды и температуры в среде обитания создают множество разных ситуаций, благоприятных и наоборот. Соотношение температуры и влажности характеризует климат конкретной территории и важно для выбора популяцией вида стации обитания. Наиболее благоприятные условия складываются в диапазоне температур 17-23°С и в диапазоне относительной влажности воздуха 85-100%.
По отношению к влажности различают эвригигробионтные и стеногигробионтные организмы. Первые живут в широком диапазоне содержания влаги, а для вторых она должна быть либо высокой, либо низкой, либо промежуточной между первыми двумя. Это относится и к растениям и к животным, несмотря на то, что вторые имеют возможность отыскивать места с оптимальной влажностью. И те и другие могут легко переносить дефицит влаги (копытные, верблюд, варан, пищуха-сеноставка, из растений: лишайники, суккуленты, многие злаки, полыни, и т.д.), а могут и вовсе не выносить сухости (земноводные, пресмыкающиеся, ластоногие, из растений: все плавающие растения, сфагновые мхи, многие папоротники, из высших: недотрога обыкновенная – быстро теряет тургор, калужницы и др.).
|
|
|
По отношению к водному режиму экотопа (экотоп – совокупность факторов местообитания) растения делятся на влаголюбивые (гидатофиты, гидрофиты, гигрофиты), сухолюбивые (ксерофиты) и умеренно влаголюбивые (мезофиты).
Гидатофиты — это водные растения, целиком или почти целиком погруженные в воду. Среди них — цветковые, которые вторично перешли к водному образу жизни (элодея, рдесты, водяные лютики, валлиснерия, уруть и др.). Вынутые из воды, эти растения быстро высыхают и погибают. У них редуцированы устьица и нет кутикулы. Листовые пластинки у гидатофитов, как правило, тонкие, без дифференцировки мезофилла, часто рассеченные, что способствует более полному использованию ослабленного в воде солнечного света и усвоению СО2. Нередко выражена разнолистность- гетерофиллия; у многих видов есть плавающие листья, имеющие световую структуру. Поддерживаемые водой побеги часто не имеют механических тканей, в них хорошо развита аэренхима.
Корневая система цветковых гидатофитов сильно редуцирована, иногда отсутствует совсем, или утратила свои основные функции (у рясок). Поглощение воды и минеральных солей происходит всей поверхностью тела. Цветоносные побеги, как правило, выносят цветки над водой (реже опыление совершается в воде), а после опыления побеги снова могут погружаться, и созревание плодов происходит под водой (валлиснерия, элодея, рдесты и др.)
Гидрофиты — это растения наземно-водные, частично погруженные в воду, растущие по берегам водоемов, на мелководьях, на болотах. Встречаются в районах с самыми разными климатическими условиями. К ним можно отнести тростник обыкновенный, частуху подорожниковую, вахту, трехлистную, калужницу болотную и другие виды. У них лучше, чем у гидатофитов, развиты проводящие и механические ткани. Хорошо выражена аэренхима. В аридных районах при сильной инсоляции их листья имеют световую структуру. У гидрофитов есть эпидермис с устьицами, интенсивность транспирации очень высока, и они могут расти только при постоянном интенсивном поглощении воды.
|
|
|
Гигрофиты (папоротник, росянка, недотрога обыкновенная, все бальзамины, рис, кислица) обитают в очень влажных местах и обладают низкой засухоустойчивостью. У них всегда открыты устьица и процесс транспирации регулируется слабо. Устьица располагаются с обеих сторон, немногочисленны. Листья крупные тонкие. Потеря 15-20% запаса воды для них невосполнима. Они растут или в глубокой тени по пологом влажного леса (теневые гигрофиты) или на открытом месте на переувлажненных или покрытых водой почвах (световые гигрофиты). Для них характерны толстые слаборазветвленные корни с минимальным количеством сосущих корней. В органах обилие воздушных полостей (аэренхима) для аэрации тканей.
Мезофиты – способны непродолжительно переносить незначительные почвенную и атмосферную засухи. К ним относятся луговые и многие лесные травы (неморальные), лиственные и хвойные деревья лесов умеренной полосы, многие кустарники, большинство сельскохозяйственных культур. Устьица расположены на нижней стороне листьев. Листья большие с умеренно развитыми тканями. Благодаря регулированию устьичной транспирации, характеризуются большой пластичностью по отношению к условиям увлажнения.
Ксерофиты – растения сухого и жаркого климата и местообитаний – пустынь, степей, саванн, в лесной зоне – растения сухих сосняков и широколиственных лесов на крутых южных склонах. Они не выносят переувлажнения, но хорошо приспособились к длительным засухам. Для них характерны два способа преодоления засухи: активное регулирование водного баланса и способность выносить сильное иссушение тканей.
У ксерофитов очень мощные корневые системы – по массе в 9-10 раз превышают надземные органы. Они или экстенсивного типа (кустарники) – длинные (10-15 м), но мало разветвленные; достигают уровня грунтовых вод, или интенсивного (злаки) – охватывают небольшой объем почвы (до 1,5 м глубины) и густо ветвятся, максимально усваивая влагу. Анатомические особенности - хорошо развита водопроводящая система, сеть жилок на листовых пластинках очень густая, позволяет быстро пополнять запасы воды, израсходованной на транспирацию.
Основные черты ксерофитности: 1. Мелкие, узкие, сильно редуцированные листовые пластинки – способствуют снижению транспирации. 2. Уменьшение (сбрасывание) листовой поверхности в наиболее сухие периоды вегетации (летний листопад). 3. Защита листьев от больших потерь влаги на транспирацию благодаря развитию мощных покровных тканей, наличие разных выростов, волосков, железок на эпидермисе («войлочное опушение» – у эдельвейсов). 4. Усиленное развитие мех. тканей листа, предотвращающих обвисание листьев при потере тургора.
Виды с наиболее выраженными перечисленными свойствами представлены склерофитами (от греч. «склеро» – твердый, жесткий; саксаул, чертополох, полыни, ковыли, молочаи и др.). Устьиц много, но они при недостатке воды закрываются. Растения могут полностью терять все листья и до 15% воды.
Другая большая группа ксерофитов – суккуленты (от лат. «суккулентус» - сочный, жирный), растут в жарком сухом климате там, где проходят кратковременные, но сильные обильные ливни. Во время дождей накапливают в листьях (агавы, молодило) или стеблях (молочаи, кактус опунция) большие запасы воды, а потом медленно ее расходуют.
Эуксерофиты – растения степей с розеточной и полурозеточной жизненной формой (кошачья лапка) и сильным опушением листьев.
Стипоксерофиты – тоже растения степных экосистем («стипо» – степь), узколистные, дерновинные злаки (вейники, типчаки, тонконог, мискантус), из с/х культур – кукуруза. Они слабо транспирируют, в сухую погоду листья сворачиваются в трубочку.
Эфемеры (весенние и осенние) – однолетние растения (незабудка песчаная, вероника весенняя, маки альпийские,), и эфемероиды – многолетние растения (крокусы, тюльпаны, прострелы), тоже обитатели засушливых местообитаний. Они избегают летних засух в связи с особенностями жизненных циклов. В короткие сроки – за 15-30 дней, растения успевают пройти весь жизненный цикл и уйти на покой до следующей весны.
Способы регуляции водного баланса у животных разнообразнее, чем у растений. Их можно разделить на поведенческие, морфологические и физиологические.
К числу поведенческих приспособлений относятся поиски водопоев, выбор мест обитания, рытье нор и т. п. В норах влажность воздуха приближается к 100%, даже когда на поверхности очень сухо. Это снижает необходимость испарения через покровы, экономит влагу в организме. Для пойкилотермных основной путь сохранения водного баланса при жизни в пустыне — это избегание излишних тепловых нагрузок
К морфологическим способам поддержания нормального водного баланса относятся образования, способствующие задержанию воды в теле: раковины наземных улиток, ороговевшие покровы рептилий, развитие эпикутикулы у насекомых и т. п.
Физиологические приспособления к регуляции водного обмена — это способность к образованию метаболической влаги, экономии воды при выделении мочи и кала, развитие выносливости к обезвоживанию, величина потоотделения
Среди животных тоже можно выделить три экологических группы, но из-за подвижного образа жизни они выражены неявно.
Гигрофилы – не могут накапливать и долго удерживать в тканях запасы воды – многие членистоногие: мокрицы, ногохвостки, комары, белоножки (гнус), а также наземные моллюски и амфибии. Нуждаются в постоянно высокой влажности воздуха. Мезофиллы – животные, обитающие в условиях умеренной влажности. Их большинство, как среди насекомых, так и среди млекопитающих. Ксерофилы – сухолюбы и термофилы одновременно, не переносят высокую влажность воздуха. У них хорошо развиты механизмы водообмена и функции удержания воды в теле. У пресмыкающихся отсутствуют кожные железы, из тела выделяется мочевая кислота, а не мочевина. У черепахи вода запасается в мочевом пузыре, грызуны воду получают с пищей. Верблюд, тушканчики, курдючные овцы воду получает в результате окисления жиров, при котором образуется метаболическая вода.
Тема 3. СРЕДЫ ОБИТАНИЯ И ИХ ХАРАКТЕРИСТИКИ
В процессе исторического развития живые организмы освоили четыре среды обитания. Первая – вода. В воде жизнь зародилась и развивалась многие миллионы лет. Вторая – наземно-воздушная – на суше и в атмосфере возникли и бурно адаптировались к новым условиям растения и животные. Постепенно преобразуя верхний слой суши - литосферы, они создали третью среду обитания – почву, а сами стали четвертой средой обитания.
Водная среда обитания
Вода покрывает 71% площади земного. Основная масса воды сосредоточена в морях и океанах – 94-98%, в полярных льдах содержится около 1,2% воды и совсем малая доля – менее 0,5%, в пресных водах рек, озер и болот.
В водной среде обитает около 150 000 видов животных и 10 000 растений, что составляет соответственно всего 7 и 8 % от общего числа видов Земли.
 В морях-океанах, как в горах, выражена вертикальная зональность. Особенно сильно различаются по экологии пелагиаль – вся толща воды, и бенталь – дно. Толща воды – пелагиаль, по вертикали делится на несколько зон: эпипелагеаль, батипелагеаль, абиссопелагиаль и ультраабиссопелагиаль (рис. 2).
В морях-океанах, как в горах, выражена вертикальная зональность. Особенно сильно различаются по экологии пелагиаль – вся толща воды, и бенталь – дно. Толща воды – пелагиаль, по вертикали делится на несколько зон: эпипелагеаль, батипелагеаль, абиссопелагиаль и ультраабиссопелагиаль (рис. 2).
В зависимости от крутизны спуска и глубины на дне тоже выделяют несколько зон, которым соответствуют указанные зоны пелагиали:
- литоральная – кромка берега, заливаемая во время приливов.
- супралиторальная – часть берега выше верхней приливной черты, куда долетают брызги прибоя.
- сублиторальная – плавное понижение суши до 200м.
- батиальная – крутое понижение суши (материковый склон),
- абиссальная – плавное понижение дна океанского ложа; глубина обеих зон вместе достигает 3-6 км.
- ультраабиссальная – глубоководные впадины от 6 до 10 км.
Экологические группы гидробионтов. Наибольшим разнообразием жизни отличаются теплые моря и океаны (40000 видов животных) в области экватора и тропиках, к северу и югу происходит обеднение флоры и фауны морей в сотни раз. Что касается распределения организмов непосредственно в море, то основная масса их сосредоточена в поверхностных слоях (эпипелагиаль) и в сублиторальной зоне. В зависимости от способа передвижения и пребывания в определенных слоях, морские обитатели подразделяются на три экологические группы: нектон, планктон и бентос.
Нектон (nektos – плавающий) - активно передвигающиеся крупные животные, способные преодолевать большие расстояния и сильные течения: рыбы, кальмары, ластоногие, киты. В пресных водоемах к нектону относятся и земноводные и множество насекомых.
Планктон (planktos – блуждающий, парящий) – совокупность растений (фитопланктон: диатомовые, зеленые и сине-зеленые (только пресные водоемы) водоросли, растительные жгутиконосцы, перидинеи и др.) и мелких животных организмов (зоопланктон: мелкие ракообразные, из более крупных – крылоногие моллюски, медузы, гребневики, некоторые черви), обитающих на разной глубине, но не способных к активным передвижениям и к противостоянию течениям. В состав планктона входят и личинки животных, образуя особую группу – нейстон. Это пассивно плавающее «временное» население самого верхнего слоя воды, представленное разными животными (десятиногие, усоногие и веслоногие ракообразные, иглокожие, полихеты, рыбы, моллюски и др.) в личиночной стадии. Выше нейстона располагается плейстон – это организмы, у которых верхняя часть тела растет над водой, а нижняя – в воде (ряска – Lemma, сифонофоры и др.). Планктон играет важную роль в трофических связях биосферы, т.к. является пищей для многих водных обитателей, в том числе основным кормом для усатых китов (Myatcoceti).
Бентос (benthos – глубина) – гидробионты дна. Представлен в основном прикрепленными или медленно передвигающимися животными (зообентос: фораминефоры, рыбы, губки, кишечнополостные, черви, моллюски, асцидии, и др.), более многочисленными на мелководье. На мелководье в бентос входят и растения (фитобентос: диатомовые, зеленые, бурые, красные водоросли, бактерии). На глубине, где нет света, фитобентос отсутствует. Наиболее богаты фитобентосом каменистые участки дна.
Высокая плотность водной среды определяет особый состав и характер изменения жизнеобеспечивающих факторов. Одни из них те же, что и на суше – тепло, свет, другие специфические: давление воды (с глубиной увеличивается на 1 атм. на каждые 10 м), содержание кислорода, состав солей, кислотность. Благодаря высокой плотности среды, значения тепла и света с градиентом высоты изменяются гораздо быстрее, чем на суше.
Тепловой режим. Для водной среды характерен меньший приход тепла, т.к. значительная часть его отражается, и не менее значительная часть расходуется на испарение. Согласуясь с динамикой наземных температур, температура воды обладает меньшими колебаниями суточных и сезонных температур. Более того, водоемы существенно выравнивают ход температур в атмосфере прибрежных районов. При отсутствии ледового панциря моря в холодное время года оказывают отепляющее действие на прилегающие территории суши, летом – охлаждающее и увлажняющее.
 Диапазон значений температуры воды в Мировом океане составляет 38° (от -2 до +36°С), в пресных водоемах – 26° (от -0,9 до +25°С). С глубиной температура воды резко падает. До 50 м наблюдаются суточные колебания температуры, до 400 – сезонные, глубже она становится постоянной, опускаясь до +1-3°С. Поскольку температурный режим в водоемах сравнительно стабилен, их обитателям свойственна стенотермность.
Диапазон значений температуры воды в Мировом океане составляет 38° (от -2 до +36°С), в пресных водоемах – 26° (от -0,9 до +25°С). С глубиной температура воды резко падает. До 50 м наблюдаются суточные колебания температуры, до 400 – сезонные, глубже она становится постоянной, опускаясь до +1-3°С. Поскольку температурный режим в водоемах сравнительно стабилен, их обитателям свойственна стенотермность.
В связи с разной степенью прогревания верхних и нижних слоев в течение года, приливами и отливами, течениями, штормами происходит постоянное перемешивание водных слоев. Роль перемешивания воды для водных обитателей исключительно велика, т.к. при этом выравнивается распределение кислорода и питательных веществ внутри водоемов, обеспечивая обменные процессы между организмами и средой.
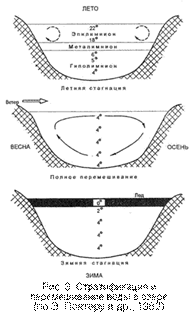
В стоячих водоемах (озерах) умеренных широт весной и осенью имеет место вертикальное перемешивание, и в эти сезоны температура во всем водоеме становится однородной, т.е. наступает гомотермия. Летом и зимой в результате резкого усиления прогревания или охлаждения верхних слоев перемешивание воды прекращается. Это явление называется температурной дихотомией, а период временного застоя – стагнацией (летней или зимней). Летом более легкие теплые слои остаются на поверхности, располагаясь над тяжелыми холодными (рис. 3). Зимой, наоборот, в придонном слое более теплая вода, так как непосредственно подо льдом температура поверхностных вод меньше +4°С и они в силу физико-химических свойств воды становятся более легкими, чем вода с температурой выше +4°С.
В периоды стагнаций четко выделяются три слоя: верхний (эпилимнион) с наиболее резкими сезонными колебаниями температуры воды, средний (металимнион или термоклин), в котором происходит резкий скачок температур, и придонный (гиполимнион), в котором температура в течение года изменяется слабо. В периоды стагнаций в толще воды образуется дефицит кислорода – летом в придонной части, а зимой и в верхней, вследствие чего в зимний период нередко происходят заморы рыбы.
Световой режим. Интенсивность света в воде сильно ослаблена из-за его отражения поверхностью и поглощения самой водой. Это сильно сказывается на развитии фотосинтезирующих растений.
Поглощение света тем сильнее, чем меньше прозрачность воды, которая зависит от количества взвешенных в ней частиц (минеральные взвеси, планктон). Уменьшается она при бурном развитии мелких организмов летом, а в умеренных и северных широтах – еще и зимой, после установления ледового покрова и укрытия его сверху снегом.
Прозрачность характеризуют предельной глубиной, на которой еще виден специально опускаемый белый диск диаметром около 20 см (диск Секки). Самые прозрачные воды — в Саргассовом море: диск виден до глубины 66,5 м. В Тихом океане диск Секки виден до 59 м, в Индийском — до 50, в мелких морях — до 5—15м. Прозрачность рек в среднем 1—1,5 м, а в самых мутных реках всего несколько сантиметров.
В океанах, где вода очень прозрачна, на глубину 140 м проникает 1% световой радиации, а в небольших озерах на глубине 2 м проникает всего лишь десятые доли процента. Лучи разных частей спектра поглощаются в воде неодинаково, вначале поглощаются красные лучи. С глубиной становится все темнее, и цвет воды становится вначале зеленым, затем голубым, синим и в конце – сине-фиолетовым, переходя в полный мрак. Соответственно меняют цвет и гидробионты, адаптирующиеся не только к составу света, но и к его недостатку – хроматическая адаптация. В светлых зонах, на мелководьях, преобладают зеленые водоросли (Chlorophyta), хлорофилл которых поглощают красные лучи, c глубиной они сменяются бурыми (Phaephyta) и далее красными (Rhodophyta). На больших глубинах фитобентос отсутствует.
К недостатку света растения приспособились развитием хроматофоров крупных размеров, а также увеличением площади ассимилирующих органов (индекса листовой поверхности). Для глубоководных водорослей типичны сильно рассеченные листья, пластинки листьев тонкие, просвечивающиеся.
Животные, как и растения, закономерно меняют свою окраску с глубиной. В верхних слоях они ярко окрашены в разные цвета, в сумеречной зоне (морской окунь, кораллы, ракообразные) окрашены в цвета с красным оттенком – удобнее скрываться от врагов. Глубоководные виды лишены пигментов. В темных глубинах океана в качестве источника зрительной информации организмы используют свет, испускаемый живыми существами- биолюминесценция.
Высокая плотность (1 г/см3, что в 800 раз больше плотности воздуха) и вязкость воды ( в 55 раз выше, чем у воздуха) привела к развитию специальных адаптаций гидробионтов:
1) У растений очень слабо развиты или вовсе отсутствуют механические ткани – им опора сама вода. Большинству свойственна плавучесть, за счет воздухоносных межклеточных полостей. Характерно активное вегетативное размножение, развитие гидрохории – вынос цветоносов над водой и распространение пыльцы, семян и спор поверхностными течениями.
2) У живущих в толще воды и активно плавающих животных тело имеет обтекаемую форму и смазано слизью, уменьшающей трение при передвижении. Развиты приспособления для повышения плавучести: скопления жира в тканях, плавательные пузыри у рыб, воздухоносные полости у сифонофор. У пассивно плавающих животных увеличивается удельная поверхность тела за счет выростов, шипов, придатков; тело уплощается, происходит редукция скелетных органов. Разные способы передвижения: изгибание тела, с помощью жгутиков, ресничек, реактивный способ передвижения (головоногие моллюски).
У придонных животных исчезает или слабо развит скелет, увеличиваются размеры тела, обычна редукция зрения, развитие осязательных органов.
Течения. Характерная черта водной среды – подвижность. Она обусловлена приливами и отливами, морскими течениями, штормами, разными уровнями высотных отметок русел рек. Приспособления гидробионтов:
1) В проточных водоемах растения прочно прикрепляются к неподвижным подводным предметам. Донная поверхность для них в первую очередь – субстрат. Это зеленые и диатомовые водоросли, водяные мхи. Мхи даже образуют плотный покров на быстрых перекатах рек. В прибойно-отливной полосе морей и многие животные имеют приспособления для прикрепления ко дну (брюхоногие моллюски, усоногие раки), или же прячутся в расщелинах.
2) У рыб проточных вод тело в поперечнике круглое, а у рыб, обитающих у дна, как и у придонных беспозвоночных животных, тело плоское. У многих на брюшной стороне есть органы фиксации к подводным предметам.
Соленость воды.
Природным водоемам свойствен определенный химический состав. Преобладают карбонаты, сульфаты, хлориды. В пресных водоемах концентрация солей не более 0,5 ‰ (причем около 80% составляют карбонаты), в морях – от 12 до 35 ‰ (в основном хлориды и сульфаты). При солености более 40 промилле водоем называют гипергалинным или пересоленным.
1) В пресной воде (гипотоническая среда) хорошо выражены процессы осморегуляции. Гидробионты вынуждены постоянно удалять проникающую в них воду, они гомойосмотичны (инфузории каждые 2-3 минуты «прокачивают» через себя количество воды, равное ее весу). В соленой воде (изотоническая среда) концентрация солей в телах и тканях гидробионтов одинакова (изотонична) с концентрацией солей, растворенных в воде – они пойкилоосмотичны. Поэтому у обитателей соленых водоемов осморегуляторные функции не развиты, и они не смогли заселить пресные водоемы.
2) Водные растения способны поглощать воду и питательные вещества из воды – «бульона», всей поверхностью, поэтому у них сильно расчленены листья и слабо развиты проводящие ткани и корни. Корни служат в основном для прикрепления к подводному субстрату. У большинства растений пресных водоемов есть корни.
Типично морские и типично пресноводные виды – стеногалинные, не переносят значительных изменений в солености воды. Эвригалинных видов немного. Они обычны в солоноватых водах (пресноводный судак, щука, лещ, кефаль, приморские лососи).
Состав газов в воде.
В воде кислород важнейший экологический фактор. В насыщенной кислородом воде содержание его не превышает 10 мл в 1 л, это в 21 раз ниже, чем в атмосфере. При перемешивании воды, особенно в проточных водоемах и при уменьшении температуры содержание кислорода возрастает. Некоторые рыбы очень чувствительны к дефициту кислорода (форель, гольян, хариус) и потому предпочитают холодные горные реки и ручьи. Другие рыбы (карась, сазан, плотва) неприхотливы к содержанию кислорода и могут жить на дне глубоких водоемов. Многие водяные насекомые, личинки комаров, легочные моллюски тоже толерантны к содержанию кислорода в воде, потому что они время от времени поднимаются к поверхности и заглатывают свежий воздух.
Углекислого газа в воде достаточно (40-50 см3/л – почти в 150 раз больше, чем в воздухе. Он используется в фотосинтезе растений и идет на формирование известковых скелетных образований животных (раковины моллюсков, покровы ракообразных, каркасы радиолярий и др.).
Кислотность. В пресноводных водоемах кислотность воды, или концентрация водородных ионов, варьирует гораздо сильнее, чем в морских – от pH=3,7-4,7 (кислые) до pH=7,8 (щелочные). Кислотностью воды определяется во многом видовой состав растений гидробионтов. В кислых водах болот растут сфагновые мхи и живут в обилии раковинные корненожки, но нет моллюсков-беззубок (Unio), редко встречаются другие моллюски. В щелочной среде развиваются многие виды рдестов, элодея. Большинство пресноводных рыб живут в диапазоне pH от 5 до 9 и массово гибнут за пределами этих значений. Максимально продуктивны воды с рН 6,5-8,5.
Кислотность морской воды убывает с глубиной.
Кислотность может служить индикатором скорости общего метаболизма сообщества. В водах с низким рН содержится мало биогенных элементов, поэтому продуктивность здесь крайне мала.
Гидростатическое давление в океане имеет большое значение. С погружением в воду на 10 м давление возрастает на 1 атмосферу. В самой глубокой части океана давление достигает 1000 атмосфер. Многие животные способны переносить резкие колебания давления, особенно, если у них в теле нет свободного воздуха. В противном случае возможно развитие газовой эмболии. Высокие давления, характерные для больших глубин, как правило, угнетают процессы жизнедеятельности.
По количеству доступного для гидробионтов органического вещества водоемы можно разделить на: - олиготрофные (голубые и прозрачные) – небогатые на корм, глубокие, холодные; - эутрофные (зеленые) – богатые кормом, теплые; дистрофные (коричневые) – бедные кормом, кислые вследствие попадания большого количества гуминовых кислот почвы.
Евтрофикация – обогащение водоёмов органическими питательными веществами под действием антропогенного фактора (н-р, спуска сточных вод).
|
|
|
