 |


Методы изучения физического развития
|
|
|
|
 Перед началом изучения данных роста важно получить представление о методах получения этих данных. Существует два основных подхода в изучении физического роста. Первый основывается на технике измерения живых организмов (включая людей), где сами измерения не наносят ущерба организму и оставляют возможность проведения повторных измерений в другое время. Второй подход включает в себя проведение экспериментов по манипуляции роста. Это подразумевает детальное изучение подопытного, которое может иметь разрушительные последствия. По этой причине такие эксперименты запрещено проводить на людях.
Перед началом изучения данных роста важно получить представление о методах получения этих данных. Существует два основных подхода в изучении физического роста. Первый основывается на технике измерения живых организмов (включая людей), где сами измерения не наносят ущерба организму и оставляют возможность проведения повторных измерений в другое время. Второй подход включает в себя проведение экспериментов по манипуляции роста. Это подразумевает детальное изучение подопытного, которое может иметь разрушительные последствия. По этой причине такие эксперименты запрещено проводить на людях.
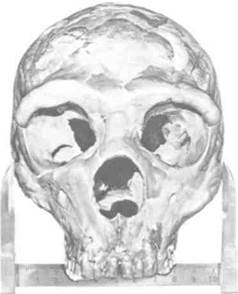
Рис. 2-9. Краниометрические исследования основаны на измерениях между ориентирами на сухих черепах, обычно находившихся среди останков первобытных людей. Классическим краниометрическим измерением является ширина отрезка между обеими скуловыми костями, как показано на снимке. Краниометрия остается важным инструментом изучения доисторических популяций.
Измерительные подходы
Первым методом измерения для изучения роста, с которого началась такая наука, как физическая антропология, является краниометрия, основанная на измерении черепов, найденных среди человеческих останков (рис. 2-9). Краниометрия изначально использовалась для изучения неандертальцев и кроманьонцев, черепа которых были найдены в пещерах Европы в XVIII и XIX веках. Из этого скелетного материала по крупицам собирались знания о вымерших популяциях и об их моделях развития при помощи сравнения черепов. Преимуществом краниометрии было то, что достаточно точные измерения производились на сухих черепах; но ее важным недостатком для изучения роста было то, что все данные о росте были кросс-секционными.
|
|
|
Кросс-секционные — это значит, что хотя в популяции присутствуют разные возрастные группы, один и тот же индивидуум может быть измерен только в одной точке времени. Также возможно проводить измерения скелета на живых людях. В данной технике, названной антропометрией, различные ориентиры на сухих черепах измеряются просто по мягким тканям, покрывающим эти костные ориентиры. Например, можно измерить длину черепа от переносицы до задней наиболее выпуклой части затылка. Это измерение может быть произведено как на высушенном черепе, так и на живом человеке, но результаты будут разными из-за толщины слоя мягкой ткани на обоих ориентирах. Хотя из-за мягких тканей возникают отклонения, антропометрия дает возможность непосредственно отслеживать развитие, производя регулярные измерения у одного человека через определенные промежутки времени. В последние годы антропометрические исследования Farkas предоставили новые данные о лицевых пропорциях человека и их изменении с течением времени1.
Третья измерительная техника, цефалометрическая рентгенология, имеет большое значение не только в исследовании роста, но и в клинической оценке состояния ортодонтических пациентов. Эта техника требует точной фиксации головы перед снимком и увеличением. Этот подход имеет преимущества как краниометрии, так и антропометрии. Он позволяет производить непосредственные измерения костей, поскольку они видны сквозь мягкие ткани на снимке (рис. 2-10), но также позволяет следить за развитием индивидуума с течением времени. Недостатком цефалометрической рентгенографии является двухгогоскостное изображение трехмерной структуры, и поэтому даже при точной фиксации головы возможны не все измерения.

Рис. 2-10. Цефалометрическая рентгенография получила название благодаря использованию устройства фиксации головы для обеспечения ее точной ориентации. Это позволяет производить сравнение внешних и внутренних измерений представителей одной группы людей или измерять одного и того же индивидуума в разное время, поскольку есть возможность повторить точное положение головы. Данный снимок сделан при естественном положении головы (NHP). Обратите внимание на цепочку, обозначающую вертикаль. В положении NHP естественная горизонталь должна быть перпендикулярна вертикальной цепочке (см. главу 6).
|
|
|
 В некоторой степени это можно преодолеть с помощью нескольких рентгеновских снимков, сделанных из разных положений и используя триангуляцию для расчета наклонных расстояний. Общая модель черепно-лицевого развития была выведена при помощи краниометрических и антропометрических исследований до изобретения цефалометрической рентгенографии, но большинство современных изображений черепно-лицевого развития основано на цефалометрических исследованиях.
В некоторой степени это можно преодолеть с помощью нескольких рентгеновских снимков, сделанных из разных положений и используя триангуляцию для расчета наклонных расстояний. Общая модель черепно-лицевого развития была выведена при помощи краниометрических и антропометрических исследований до изобретения цефалометрической рентгенографии, но большинство современных изображений черепно-лицевого развития основано на цефалометрических исследованиях.
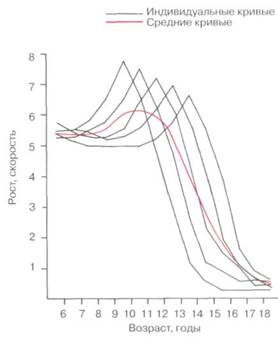
Рис. 2-11. При изображении в виде графика на хронологической шкале данных скорости роста группы индивидуумов с различными сроками подросткового скачка в развитии становится ясным, что средняя кривая не отражает ни одну модель роста конкретного индивидуума. Такое сглаживание индивидуальных вариаций характерно для статичных данных и ограничивает использование статичного метода для изучения роста. Только при исследовании индивидуумов с течением времени в рамках динамичного исследования представляется возможным увидеть детали модели роста.
Как антропометрические, так и цефалометрические данные могут быть выражены лишь в статичном, а не динамичном аспекте. Очевидным является то, что гораздо проще и быстрее производить статичные исследования, собирая данные по нескольким индивидуумам различных возрастов, чем тратить многие годы на изучение регулярных измерений одного человека. По этой причине большинство исследований статичны. Однако при использовании данного подхода за вариативностью могут скрываться особенности модели роста, в частности, когда отсутствует коррекция отклонения по срокам (рис. 2-11). Скачки кривой роста, которые могут происходить почти у каждого индивидуума, могут наблюдаться при статичном исследовании, только если они совпадают по времени у каждого исследуемого, что маловероятно. Динамичные исследования эффективны в том смысле, что большой объем информации может быть получен при исследовании относительно небольшого числа субъектов, меньшего, чем требовалось бы для статичного исследования. Кроме того, динамичные данные отражают индивидуальные отклонения, то есть отклонения, произошедшие под влиянием времени.
|
|
|
 Данные измерений могут быть графически представлены разными способами, и часто появляется возможность проследить изменения в развитии, варьируя методы их отражения. Например, мы уже видели, что данные роста могут быть представлены в виде изображения функции возраста, что называется «дистанционная» кривая или кривая «скорости», отражающая не общую длину, а ежегодный прирост (см. рис. 2-6).
Данные измерений могут быть графически представлены разными способами, и часто появляется возможность проследить изменения в развитии, варьируя методы их отражения. Например, мы уже видели, что данные роста могут быть представлены в виде изображения функции возраста, что называется «дистанционная» кривая или кривая «скорости», отражающая не общую длину, а ежегодный прирост (см. рис. 2-6).
Рис. 2-12. Данные увеличения массы раннего эмбриона; необработанные данные изображены в виде графика черного цвета, а те же данные после логарифмической трансформации — в виде графика красного цвета. На данном этапе происходит резкое увеличение массы эмбриона, но прямая линия графика после трансформации свидетельствует о том, что степень деления клеток остается практически постоянной. При большем числе клеток происходит больше делений и масса увеличивается быстрее. (Цит. по: Lowery GH: Growth and development of children, ed 8, Chicago, 1986, Mosby.)
 Изменения в степени роста легче проследить на кривой скорости, чем на дистанционной кривой. Различные другие математические трансформации могут быть применены к данным роста для облегчения их понимания. Например, прибавление в массе эмбриона на ранней стадии идет по логарифмической или экспоненциальной кривой, потому что развитие базируется на делении клеток: чем больше клеток, тем больше происходит делений. Если график строится на основе одних и тех же данных с использованием логарифма массы, получается прямолинейный график (рис. 2-12). Это указывает на то, что степень размножения клеток эмбриона остается более или менее постоянной.
Изменения в степени роста легче проследить на кривой скорости, чем на дистанционной кривой. Различные другие математические трансформации могут быть применены к данным роста для облегчения их понимания. Например, прибавление в массе эмбриона на ранней стадии идет по логарифмической или экспоненциальной кривой, потому что развитие базируется на делении клеток: чем больше клеток, тем больше происходит делений. Если график строится на основе одних и тех же данных с использованием логарифма массы, получается прямолинейный график (рис. 2-12). Это указывает на то, что степень размножения клеток эмбриона остается более или менее постоянной.
|
|
|
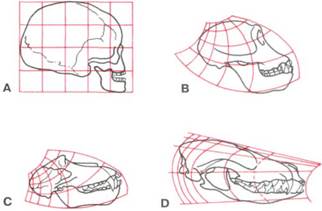
Рис. 2-13. В начале 1900-х годов D'Arcy Thompson продемонстрировал математическую трансформацию сетки лицевой модели от человека (А) до шимпанзе (В), обезьяны (С), собаки (D) или других животных. Использование данного метода выявило доселе неизвестные сходства между различными видами. (Цит. по: Thompson JT: On growth and form, Cambridge, Mass., 1971, Cambridge University Press.)
Более сложные математические трансформации были использованы много лет назад D'Arcy Thompson2 для выявления схожести пропорций и изменений роста, которых ранее не наблюдалось (рис. 2-13).
Математические трансформации также применяются к изменениям роста на цефалометрических рентгенограммах в попытке определения модели изменений, которая не может быть определена другим способом. Для правильной интерпретации данных после математической трансформации важно понимать, каким образом данные были трансформированы, однако метод играет важную роль в разъяснении концепции развития.
Экспериментальные подходы
 Много новых знаний о развитии скелета было получено при использовании техники прижизненного окрашивания, при котором вещества, окрашивающие минерализованные ткани (или иногда мягкие ткани) инъецировались в тело животного. Эти красители оставались в костях и зубах и могли быть найдены после смерти животного. Данный метод был разработан в XVlII веке великим английским анатомом John Hunter. Hunter заметил, что кости свиней, в корме которых иногда попадались текстильные отходы, были окрашены интересным образом. Он открыл, что активным агентом был краситель под названием ализарин, до сих пор использующийся в исследованиях при помощи прижизненного окрашивания (рис. 2-14).
Много новых знаний о развитии скелета было получено при использовании техники прижизненного окрашивания, при котором вещества, окрашивающие минерализованные ткани (или иногда мягкие ткани) инъецировались в тело животного. Эти красители оставались в костях и зубах и могли быть найдены после смерти животного. Данный метод был разработан в XVlII веке великим английским анатомом John Hunter. Hunter заметил, что кости свиней, в корме которых иногда попадались текстильные отходы, были окрашены интересным образом. Он открыл, что активным агентом был краситель под названием ализарин, до сих пор использующийся в исследованиях при помощи прижизненного окрашивания (рис. 2-14).
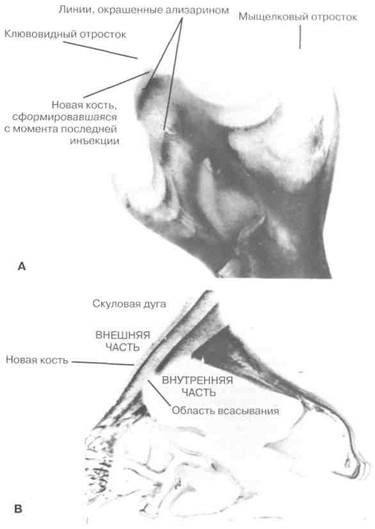
Рис. 2-14. А — нижняя челюсть крысы, получившей три инъекции ализарина с интервалом в 3 нед., и умершей через 2 нед. после последней инъекции. Реконструкция кости в ходе роста затемняет некоторые инъекционные линии, но последовательные линии в мыщелковом процессе видны довольно ясно. В — отрезок скуловой дуги того же животного, что и на снимке А. Скуловая дуга растет наружу с аппозицией кости на внешней поверхности и удалением с внутренней поверхности. Разрывы окрашенных линий на внутренней поверхности обозначают места, где кость была удалена. То, что было внешней поверхностью скуловой дуги, затем становится внутренней поверхностью и некоторое время спустя удаляется.
 Ализарин вступает в сильную реакцию с кальцием в местах обызвествления костей. Поскольку эти места претерпевают активное скелетное развитие, окрашенные метки указывают степень развития на момент инъекции. Кость быстро восстанавливается, и области, где произошло удаление кости, могут быть определены по отсутствию окрашенного материала в этих местах. В настоящее время появилась возможность проводить высокоточные исследования костных изменений в ходе черепно-лицевого развития у подопытных животных3.
Ализарин вступает в сильную реакцию с кальцием в местах обызвествления костей. Поскольку эти места претерпевают активное скелетное развитие, окрашенные метки указывают степень развития на момент инъекции. Кость быстро восстанавливается, и области, где произошло удаление кости, могут быть определены по отсутствию окрашенного материала в этих местах. В настоящее время появилась возможность проводить высокоточные исследования костных изменений в ходе черепно-лицевого развития у подопытных животных3.
|
|
|
Рис. 2-15. Тетрациклиновое окрашивание зубов у мальчика, получившего большую дозу тетрациклина при лечении инфекционного заболевания верхних дыхательных путей в раннем детстве. Исходя из распространения окрашенности, очевидно, что тетрациклин не прописывался в грудном возрасте, но давался в больших дозах уже к моменту формирования коронок центральных резцов или приблизительно в возрасте 2,5 лет.
Хотя исследования с использованием прижизненного окрашивания невозможно проводить на людях, прижизненное окрашивание все же может встречаться. Многие дети, рожденные в конце 1950-х и начале 1960-х годов, проходили курс лечения инфекционных заболеваний антибиотиком тетрациклином. Слишком поздно было обнаружено, что тетрациклин является отличным прижизненным красителем, который связывается кальцием в местах роста, как и ализарин. Нарушение цвета резцов в результате лечения тетрациклином во время минерализации зубов стало для некоторых эстетической катастрофой (рис. 2-15), и поэтому теперь для лечения инфекций у детей тетрациклин применяется очень редко.
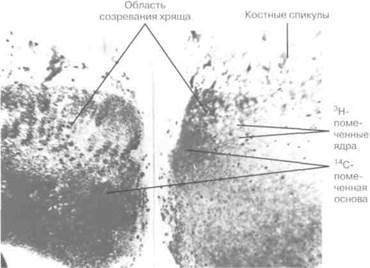 Рис. 2-16. Результаты авторадиографии костей зародыша крысы, развивающегося с изотопами ,4С-пролин и 3Н-тимидин, имплантированными в культурную среду. Тимидин имплантирован в ДНК, которая воспроизводится при делении клетки, и помеченные ядра относятся к клеткам, претерпевшим культурный митоз. Поскольку пролин является основной составляющей коллагена, метки цитоплазмы определяют области, где был имплантирован пролин, изначально в межклеточный коллаген. А — нормальный рост в данной среде с множеством помеченных клеток и большим распространением пролина. В — замедленный рост кости с малым количеством бактериального эндотоксина.
Рис. 2-16. Результаты авторадиографии костей зародыша крысы, развивающегося с изотопами ,4С-пролин и 3Н-тимидин, имплантированными в культурную среду. Тимидин имплантирован в ДНК, которая воспроизводится при делении клетки, и помеченные ядра относятся к клеткам, претерпевшим культурный митоз. Поскольку пролин является основной составляющей коллагена, метки цитоплазмы определяют области, где был имплантирован пролин, изначально в межклеточный коллаген. А — нормальный рост в данной среде с множеством помеченных клеток и большим распространением пролина. В — замедленный рост кости с малым количеством бактериального эндотоксина.
 С развитием радиоактивных индикаторов стало возможным использование практически любого метаболита с радиоактивной окраской, вводимого в ткани в качестве прижизненного красителя. Его расположение должно, безусловно, определяться слабой радиоактивностью в месте имплантации. Гамма-излучающий изотоп "111Tc может быть использован для индикации областей быстрого роста костей человека, однако данные изображения больше подходят для диагностики локальных проблем роста (см. главу 21), чем для изучения моделей роста. В большинстве исследований роста радиоактивно помеченные материалы в тканях подопытных животных распознаются при помощи техники авторадиографии, где пленочная эмульсия накладывается на тонкие участки ткани, содержащей изотопы, а затем в темноте подвергается радиоактивному облучению. После проявления пленки наличие радиации, обозначающей области роста, может быть отмечено при рассмотрении сектора ткани через пленку (рис. 2-16).
С развитием радиоактивных индикаторов стало возможным использование практически любого метаболита с радиоактивной окраской, вводимого в ткани в качестве прижизненного красителя. Его расположение должно, безусловно, определяться слабой радиоактивностью в месте имплантации. Гамма-излучающий изотоп "111Tc может быть использован для индикации областей быстрого роста костей человека, однако данные изображения больше подходят для диагностики локальных проблем роста (см. главу 21), чем для изучения моделей роста. В большинстве исследований роста радиоактивно помеченные материалы в тканях подопытных животных распознаются при помощи техники авторадиографии, где пленочная эмульсия накладывается на тонкие участки ткани, содержащей изотопы, а затем в темноте подвергается радиоактивному облучению. После проявления пленки наличие радиации, обозначающей области роста, может быть отмечено при рассмотрении сектора ткани через пленку (рис. 2-16).
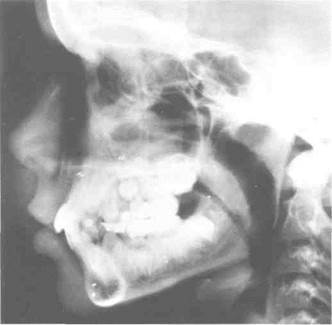
Рис. 2-17. Цефалометрическая рентгенограмма, на которой видны имплантаты, установленные перед началом лечения по изменению положения челюстей. Металлические имплантаты видны в виде небольших белых линий. Сопоставление рентгенограмм состояния до и после лечения при помощи данных имплантатов позволяет точно определить перемещения зубов и челюстей. (Снимок предоставлен Professor Beni Solow.)
Другим экспериментальным методом, применяемым для исследования роста человека, является имплантационная рентгенография. В данной технике в кости скелета, включая челюсти и кости лица, помещаются инертные металлические штифты. Эти штифты хорошо срастаются с костями без каких-либо осложнений (рис. 2-17). При помощи вживления металлических имплантатов значительно улучшается точность динамического цефалометрического анализа модели роста. Данный метод исследования, широко использованный профессором Arne Bjork и его коллегами из Королевского стоматологического колледжа в Копенгагене, Дания6, помог получить новые важные данные о моделях развития челюстей. Металлические штифты остаются в месте имплантации внутри кости при отсутствии инфекции или воспаления, что случается крайне редко. Сопоставление цефалометрических рентгенограмм с имплантированными штифтами обеспечивает точное изучение как изменений положения одной кости относительно другой, так и изменений внешнего контура костей. До проведения рентгенографических исследований с использованием имплантатов степень реконструкционных изменений контуров челюстных костей недооценивалась, а также не принималась во внимание ротационная модель челюстного развития, описанная в главе 4. Точная оценка челюстно-лицевого развития у человека до настоящего времени наилучшим образом осуществляется при помощи имплантационной рентгенографии7.
Природа развития скелета
На клеточном уровне существует только три возможности роста. Первая состоит в увеличении размеров отдельных клеток, что называют гипертрофией. Второй возможностью является увеличение числа клеток, или гиперплазия. Третья возможность — это выделение метками внеклеточного вещества, что способствует увеличению в размерах независимо от количества или размера самих клеток.
В действительности при росте скелета наблюдаются все три процесса. Гиперплазия является самой известной формой всех типов роста. Гипертрофия происходит при особых обстоятельствах, но в большинстве случаев представляет собой менее значимый механизм, чем гиперплазия. Хотя во всех тканях организма происходит выделение межклеточного вещества, это явление особенно важно при развитии скелетной системы, где происходит последующая минерализация межклеточного вещества.
Тот факт, что межклеточное вещество скелета минерализуется, ведет к важному разделению между ростом мягких, неминерализованных тканей тела и твердых, или подвергнутых кальцинозу, тканей. К твердым тканям относятся кости, зубы и некоторые хрящи. Все остальные ткани — мягкие. В большинстве случаев хрящи, задействованные в росте, ведут себя как мягкие ткани и должны быть отнесены в разряд скорее мягких, чем твердых тканей.
Рост мягких тканей происходит за счет комбинации гиперплазии и гипертрофии. Эти процессы происходят по всей площади тканей, и результат получил название интерстициального роста, что означает лишь, что данный процесс происходит в каждой точке ткани. Выделение внеклеточного вещества может также сопровождать интерстициальное развитие, однако гиперплазия и гипертрофия играют здесь соответственно первую и вторую роли. Интерстициальный рост характерен почти для всех мягких тканей и для не подверженных кальцинозу хрящей скелетной системы.
Напротив, при наличии минерализации, когда происходит процесс формирования твердых тканей, интерстициальный рост невозможен. Гиперплазия, гипертрофия и выделение межклеточного вещества все же возможны, но могут происходить лишь на поверхности, а не внутри минерализованной массы. Непосредственное добавление новой кости на поверхности существующей может происходить и происходит благодаря деятельности клеток надкостницы — мягкой мембранной ткани, покрывающей кости. В надкостнице происходит формирование новых клеток, а выделяемое межклеточное вещество минерализуется и превращается в новую кость. Данный процесс называется непосредственной или поверхностной оппозицией кости.
Интерстициальный рост является важным аспектом общего развития скелета, поскольку большая часть скелетной системы изначально моделируется в хрящах. Здесь имеется в виду основная часть черепа, а также туловища и конечностей.
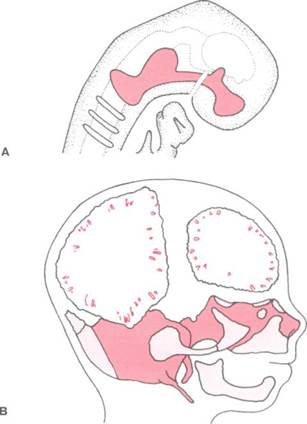 На рисунке 2-18 изображен хрящевой череп на 8-й и 12-й неделях внутриутробного развития. Начало хрящевого скелетного развития происходит на 3-м месяце внутриутробной жизни. Непрерывная хрящевая пластина проходит от носовой капсулы назад к отверстию головчатой кости у основания черепа. Следует помнить, что хрящ является практически бессосудной тканью, внутренние клетки которой снабжаются посредством диффузии через внешние слои. Безусловно, это означает, что хрящ должен быть тонким. На ранних стадиях развития крайне малый размер эмбриона позволяет скелету образоваться из гиалинового хряща, однако в ходе дальнейшего роста такое строение невозможно без внутреннего кровоснабжения.
На рисунке 2-18 изображен хрящевой череп на 8-й и 12-й неделях внутриутробного развития. Начало хрящевого скелетного развития происходит на 3-м месяце внутриутробной жизни. Непрерывная хрящевая пластина проходит от носовой капсулы назад к отверстию головчатой кости у основания черепа. Следует помнить, что хрящ является практически бессосудной тканью, внутренние клетки которой снабжаются посредством диффузии через внешние слои. Безусловно, это означает, что хрящ должен быть тонким. На ранних стадиях развития крайне малый размер эмбриона позволяет скелету образоваться из гиалинового хряща, однако в ходе дальнейшего роста такое строение невозможно без внутреннего кровоснабжения.
Рис. 2-18. Развитие и созревание хрящевого черепа (хрящ — красный цвет, кость — красная штриховка). А — схематичное изображение черепа в возрасте 8 нед. Обратите внимание на значительный участок хрящевой ткани от носовой капсулы до затылочной области. В — скелетное развитие в возрасте 12 нед. В хрящевой структуре появились центры окостенения, а также началось формирование межмембранной кости челюстей и черепной коробки. С этого момента начинается быстрая замена хрящевой ткани черепа на костную и, таким образом, остаются лишь небольшие синхондрозы, соединяющие кости основания черепа.
 В течение 4-го месяца внутриутробного развития в различных точках хрящевого черепа (а также в других частях первоначально хрящевого скелета) происходит прорастание внутрь кровеносных сосудов. Данные участки становятся центрами окостенения, в которых хрящ трансформируется в кость, и в общей хрящевой массе появляются островки кости (рис. 2-18, В). Продолжается быстрый рост хрящей, которые с такой же скоростью заменяются костью. В результате происходит быстрое увеличение относительного количества кости, а относительное (но не абсолютное) количество хряща уменьшается. В конечном итоге прежний хрящевой череп представлен лишь небольшими участками хряща, находящимися между большими областями кости, которая принимает характерные формы этмоида, клиновидной кости и затылочных костей. Рост данных хрящевых соединений между костями скелета схож с ростом в конечностях.
В течение 4-го месяца внутриутробного развития в различных точках хрящевого черепа (а также в других частях первоначально хрящевого скелета) происходит прорастание внутрь кровеносных сосудов. Данные участки становятся центрами окостенения, в которых хрящ трансформируется в кость, и в общей хрящевой массе появляются островки кости (рис. 2-18, В). Продолжается быстрый рост хрящей, которые с такой же скоростью заменяются костью. В результате происходит быстрое увеличение относительного количества кости, а относительное (но не абсолютное) количество хряща уменьшается. В конечном итоге прежний хрящевой череп представлен лишь небольшими участками хряща, находящимися между большими областями кости, которая принимает характерные формы этмоида, клиновидной кости и затылочных костей. Рост данных хрящевых соединений между костями скелета схож с ростом в конечностях.
Рис. 2-19. Эндохондральное окостенение на эпифизарной пластине. Развитие происходит при разрастании хряща в верхней части. Созревающие клетки хряща отодвинуты от области разрастания, подвергаются гипертрофии, дегенерируют и заменяются костными спикулами, как показано в нижней части снимка.
В длинных костях конечностей области окостенения возникают в центре кости и на ее концах, образуя центральное тело, называемое диафизом, и костные наконечники с каждой стороны, называемые эпифизом. Остающаяся между эпифизом и диафизом область не подверженного кальцинозу хряща называется эпифизарной пластиной (рис. 2-19). Эпифизарная хрящевая пластина длинных костей является основным центром их роста, и в действительности этот хрящ отвечает за развитие по всей длине данных костей. Надкостница на поверхности костей также имеет большое значение в увеличении толщины и изменении формы внешних контуров.
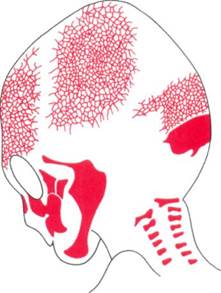 У внешнего конца каждой эпифизарной пластины расположена область активного деления хрящевых клеток. Некоторые из них, продвигаемые к диафизу в результате активности клеток, претерпевают гипертрофию, выделяют межклеточную основу и в результате дегенерируют, когда основа начинает минерализовываться, а затем быстро заменяются костной тканью (рис. 2-19). Пока степень разрастания хрящевых клеток равна или превышает скорость их созревания, рост будет продолжаться. Однако ближе к завершению нормального периода роста скорость созревания превышает скорость разрастания, последние хрящевые участки заменяются костью и эпифизарная пластина пропадает. На данном этапе развитие кости завершено, за исключением поверхностных изменений толщины, осуществляемых надкостницей.
У внешнего конца каждой эпифизарной пластины расположена область активного деления хрящевых клеток. Некоторые из них, продвигаемые к диафизу в результате активности клеток, претерпевают гипертрофию, выделяют межклеточную основу и в результате дегенерируют, когда основа начинает минерализовываться, а затем быстро заменяются костной тканью (рис. 2-19). Пока степень разрастания хрящевых клеток равна или превышает скорость их созревания, рост будет продолжаться. Однако ближе к завершению нормального периода роста скорость созревания превышает скорость разрастания, последние хрящевые участки заменяются костью и эпифизарная пластина пропадает. На данном этапе развитие кости завершено, за исключением поверхностных изменений толщины, осуществляемых надкостницей.
Рис. 2-20. Кости черепа 12-недельного зародыша, нарисованные по окрашенному ализарином образцу. (Цит. по: Langman J: Medical embryology, et 4, Baltimore, 1984, Williams & Wilkins.)
Не все кости взрослого скелета были представлены в хрящевой модели эмбриона, и кости могут осуществлять выделение костной основы непосредственно внутри соединительных тканей, без формирования промежуточных хрящей. Формирование костей по данному типу называется межмембранным костным формированием. Данный тип окостенения характерен для черепного свода и обеих челюстей (рис. 2-20).
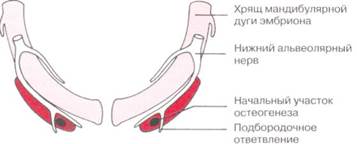
Рис. 2-21. Схематичное изображение формирования начальной кости нижней челюсти по отношению к хрящу мандибулярной дуги эмбриона и нижнему альвеолярному нерву. Формирование кости начинается сбоку от хряща и распространяется назад вдоль него без непосредственной замены хряща новой костью нижней челюсти. (Цит. по: Ten Cate AR: Oral histology, St. Louis, 1985, Mosby.)
В начале эмбрионального периода нижняя челюсть высших животных развивается в той же области, что и хрящ первой фарингеальной арки, хрящ мандибулярной дуги эмбриона. Очевидно, что нижняя челюсть является костным заменителем данного хряща, так же как и клиновидная кость заменяет хрящ в этой области.
В действительности развитие нижней челюсти начинается с конденсации мезенхимы по бокам хряща мандибулярной дуги эмбриона и продолжается в виде межмембранного костного формирования (рис. 2-21). Хрящ мандибулярной дуги эмбриона распадается и полностью исчезает с развитием кости нижней челюсти. Остатки этого хряща трансформируются в части двух небольших костей, формирующих связующие мелкие кости среднего уха, на неосновную часть нижней челюсти. Ее перихондриум существует в виде клиновидно-нижнечелюстной связки. Мыщелковый хрящ изначально развивается в виде независимого вторичного хряща, отделенного значительным промежутком от тела нижней челюсти (рис. 2-22). В начале жизни зародыша он сращивается с развивающимся ответвлением нижней челюсти.
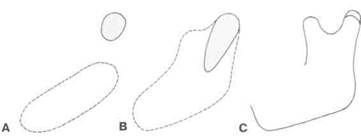
Рис. 2-22. Мыщелковый хрящ (розовый цвет) сначала развивается отдельно от тела нижней челюсти и лишь позже становится ее частью. А — отдельные области мезенхимной конденсации в возрасте 8 нед. В - слияние хряща с телом нижней челюсти в возрасте 4 мес. С - ситуация на момент рождения (в уменьшенном масштабе).
Верхняя челюсть начинает свое формирование из центра мезенхимной конденсации. Данная область расположена на боковой поверхности носовой капсулы, наиболее передней части хрящевого черепа, хотя рост хряща и способствует лишь удлинению головы и переднему перемещению верхней челюсти, но не способствует непосредственному формированию верхнечелюстной кости. Дополнительный или скуловой хрящ, формирующий процесс скулового развития, исчезает и полностью заменяется костью перед рождением, в отличие от нижнечелюстного мыщелкового хряща, который сохраняется.
В каком бы месте ни происходило межмембранное формирование кости, рост внутри минерализованной массы невозможен и кость должна формироваться исключительно аппозицией новых костей на свободной поверхности. Ее форма может изменяться посредством удаления (резорбции) кости в одной области и добавления (аппозиции) кости в другой (см. рис. 2-13). Данное равновесие аппозиции и резорбции, когда в некоторых областях происходит формирование новой кости, а в других удаление старой, является определяющим компонентом процесса роста. Реконструкция данного типа наблюдается на поверхности костей, растущих в первую очередь посредством внутрихрящевой замены, а также в костях, формируемых непосредственно внутри мембраны связующей ткани.
|
|
|
