 |


Условный рефлекс — одна из форм научения. Методы изучения высшей нервной деятельности
|
|
|
|
Научение — это выработка в онтогенезе приспособительных форм поведения организма, в том числе и навыков физического труда. Условный рефлекс — одна из форм научения (это выработанная в онтогенезе реакция организма на раздражительность, ранее индифферентной для этой реакции).
Выработка условного рефлекса. Основные правила выработки условных рефлексов:
1) условный рефлекс образуется на базе безусловного (врожденного) рефлекса (рис. 94);
2) условный стимул должен предшествовать безусловному;
3) необходимо неоднократное совпадение во времени действия индифферентного раздражителя с безусловным.
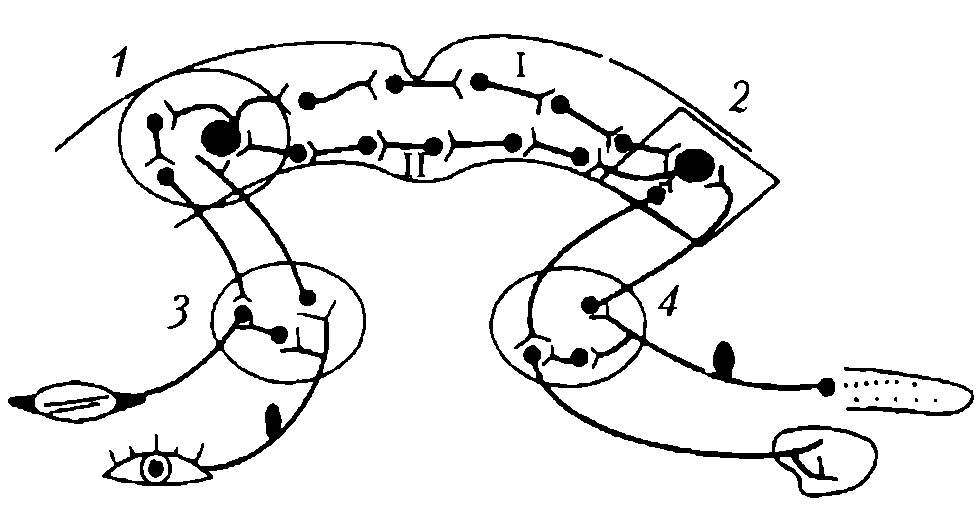
Рис. 94. Дуга условного рефлекса с двусторонней связью (по Э.А. Асратяну): 1 – корковый центр мигательного рефлекса; 2 – корковый центр пищевого рефлекса; 3,4 – подкорковые центры мигательного и пищевого рефлексов; I – прямая временная связь; II – обратная временная связь
Факторы, способствующие образованию условных рефлексов: 1) активное функциональное состояние нервного центра безусловного рефлекса (для пищевых рефлексов — наличие пищевой потребности);
2) оптимальное соотношение силы условного и безусловного раздражителей (А. Ф. Белов);
3) отсутствие значимых посторонних сигналов из внешней и внутренней среды организма;
4) оптимальное состояние организма (здоровье), в первую очередь состояние коры большого мозга.
Основные периоды формирования условного рефлекса:
1) скрытый период (условные ответы не возникают, несмотря на неоднократные сочетания условного и безусловного раздражителей);
2) период генерализации — условные реакции возникают на все посторонние сигналы вследствие иррадиации возбуждения в коре;
3) период специализации (условный ответ возникает только на тот условный сигнал, на который вырабатывался условный рефлекс), чему способствует формирование доминантного очага возбуждения в центре условного рефлекса.
|
|
|
Механизм формирования условных рефлексов. Временная связь, согласно И. П. Павлову (1903), образуется между корковыми центрами безусловного и условного раздражителей. Импульсы, вызываемые индифферентным (условным) сигналом с любых рецепторов, по нервам поступают в кору большого мозга и обеспечивают там образование очага возбуждения. Если после индифферентного сигнала дать, например, пищевое подкрепление, то возникает более мощный второй очаг возбуждения в коре большого мозга, к которому направляется ранее возникшее ииррадиирующее по коре возбуждение. Это представление позже получило подтверждение в учении о доминанте А.А. Ухтомского. На стадии сигнализации условного рефлекса важную роль играет доминанта.
Неоднократное сочетание в опытах индифферентного (условного) сигнала и безусловного раздражителя облегчает прохождение импульсов от коркового центра индифферентного сигнала к корковому представительству безусловного рефлекса (проторение пути).
Индифферентный сигнал не может быть вообще индифферентным (Э. А. Асратян), так как в начале своего применения вызывает собственную реакцию в виде ориентировочно-исследовательского рефлекса, который по мере повторения ослабляется. Замыкание происходит между центрами двух безусловных рефлексов — сигнального и подкрепляющего. Это нашло подтверждение в опытах, в которых изучали два безусловных рефлекса — мигательный, вызываемый подачей струи воздуха в глаз, и безусловный пищевой. При их сочетании выработался условный рефлекс, причем если подавали воздушную струю, то возникал пищевой рефлекс, а при подаче пищевого раздражителя отмечалось мигание.
|
|
|
Таким образом, в основе механизма выработки условного рефлекса лежат явления иррадиации процесса возбуждения, синаптической потенциации (проторение пути) и доминанта.
Временная связь замыкается с помощью механизмов памяти (период иррадиации возбуждения — электрофизиологическая память, далее — нейрохимическая, ультраструктурная и макромолекулярная память).
Компоненты условного рефлекса включают таковые безусловного, на базе которого он выработан. На начальных этапах образования условного рефлекса в период изолированного действия условного сигнала отмечается определенная очередность появления компонентов: вначале обнаруживается сопряженный компонент (дыхательный, сосудистый, сердечный и др.), имеющий значение для энергетического обеспечения последующей специфической деятельности (например, потребление пищи), затем появляется специфический поведенческий (соматический) компонент — приближение животного к пище. Позже включаются секреторные и моторные проявления ЖКТ.
Сравнительная характеристика условных и безусловных рефлексов:
1) условные рефлексы — индивидуальные (приобретаются в течение жизни), безусловные — врожденные (отражают видовые особенности организма);
2) условные рефлексы осуществляются с помощью функционально организующихся временных (замыкательных) связей, а безусловные — по анатомическим путям, определенным генетически;
3) условные рефлексы изменчивы (исчезают, когда они не нужны), а безусловные — относительно постоянны в течение жизни особи;
4) условные рефлексы для своего образования и реализации требуют целостности коры большого мозга, особенно у высших млекопитающих, а безусловные рефлексы могут осуществляться с помощью любого уровня ЦНС;
5) условные рефлексы могут образовываться с любого рецептивного поля, на самые разнообразные раздражители, а каждый безусловный рефлекс имеет свое специфическое рецептивное поле и свои специфические раздражители;
6) условные рефлексы возникают заблаговременно (готовят организм к предстоящим событиям), а безусловные рефлексы возникают в ответ на действие наличного раздражителя, которого уже нельзя избежать;
7) набор условных рефлексов безграничен, а число определенных видов безусловных рефлексов определено генетически.
|
|
|
3.1.13. Методы исследования ВНД
Общая характеристика основных методов изучения ВНД. Высшая нервная деятельность изучается с помощью выработки условных рефлексов и других видов научения и воздействия на ЦНС или исследования ее состояния.
Выработка различных условных рефлексов и других форм научения на фоне холодового выключения, разрушения и раздражения отдельных структур мозга, что позволяет выявить роль отдельных структур в замыкании временной связи. Преимущество холодового выключения заключается в том, что возможно многократно обратимо выключать тот или иной центр.
Научение при введении агонистов (ацетилхолина, адреналина, серотонина и др.), антагонистов (блокаторов); на фоне действия молекул ДНК, РНК.
Регистрация в процессе научения ЭЭГ, ВП, нейронной активности различных отделов коры большого мозга, подкорковых структур, отдельных нейронов на разной стадии выработки условного рефлекса и других видов научения, любой психической деятельности.
Исследования изменений в синаптических структурах нейронов, их отростков с помощью электронной микроскопии после выработки условных рефлексов и формирование других видов научения. В связи с компьютеризацией научных исследований весьма важным методом стала ЭЭГ.
Электроэнцефалография — это регистрация суммарного электрического поля мозга с поверхности головы, создаваемого ПД, ВПСП и ТПСП.
ЭЭГ — кривая, зарегистрированная при этом. Запись ЭЭГ с коры головного мозга называется электрокортикограммой. Регистрация ЭЭГ производится с помощью биполярных или униполярных электродов, накладываемых на кожу головы.
При этом регистрируется четыре основных ритма: α-, β-, θ- и δ-ритмы.
Альфа-ритм (8—13 Гц, до 70 мкВ) наблюдается у человека в состоянии физического и психического покоя, преобладает у 85—95% здоровых людей старше девятилетнего возраста. Лучше всего α-ритм выражен в затылочных областях мозга, в центральной и лобной областях, часто сочетается с β-ритмом. Вариантом α-ритма являются «веретена сна» длительностью 2—8 с (п. 18.4). Альфа-ритм является упорядоченным регулярным ритмом. Если он доминирует, ЭЭГ рассматривается как синхронизированная.
|
|
|
Бета-ритм имеет нерегулярную частоту (14—30 Гц), низкую амплитуду — до 30 мкВ, сменяет α-ритм при сенсорной стимуляции (десинхронизация ЭЭГ), например, при действии звука, при эмоциональном возбуждении. Наиболее выражен β-ритм в лобных, центральных областях головного мозга. Бета-ритм отражает высокий уровень функциональной активности головного мозга.
Тета-ритм (4—7 Гц, до 200 мкВ) у бодрствующего человека регистрируется обычно в передних областях мозга при длительном эмоциональном напряжении, почти всегда регистрируется в процессе развития фаз медленноволнового сна. Отчетливо регистрируется у детей, пребывающих в состоянии эмоции неудовольствия.
Дельта-ритм (0,5—3,0 Гц, 200 — 300 мкВ) регистрируется эпизодически во всех областях головного мозга при снижении функциональной активности мозга.
Также выделяют и другие ритмы ЭЭГ (таб. 12).
Таблица 12. Сравнение видов фоновой электрической активности (электроэнцефалографических ритмов) головного мозга (Kryger, 2011)
| Название ритма | Частота и амплитуда ритма | Преимущественная локализация электрической активности (локализация ритма) | Наиболее вероятные проявления в норме | Наиболее вероятные проявления при патологии |
| Дельта-ритм, σ | 0,5-4 Гц | У взрослых в лобных областях головного мозга, у детей в затылочных областях головного мозга | - У взрослых во время глубокого медленного сна - У детей - Вовремя решения некоторых задач, требующих длительного вовлечения внимания | - Повреждение подкорковых структур головного мозга - Диффузные повреждения головного мозга - Метаболическая энцефалопатия гидроцефалов - Повреждения глубоких срединных структур головного мозга |
| Тета-ритм, θ | 4-8 Гц | Проявляется в областях головного мозга, не относящихся к решению задач, связанных с движениями рук | - Дети младшего возраста - Дремота или пробуждение от сна детей старшего возраста и взрослых - В отсутствии деятельности Связаны с торможением электрических вызванных ответов головного мозга (попытки сознательного подавления испытуемым ответа на стимулы) | - Ограниченные Повреждения подкорковых структур головного мозга - Метаболическая энцефалопатия - Нарушения глубоких структур головного мозга - Иногда у гидроцефалов |
| Альфа-ритм, α | 8-13 Гц | С отведений затылочных областей головного мозга, с обеих сторон, амплитуда волн выше с недоминантной стороны. В покое – с отведений центральных областей (с3-с4) | - В расслабленном состоянии, зеркально - Глаза закрыты - Могут быть связаны с тормозным управлением в различных областях головного мозга | Кома |
| Бета-ритм, β | 13-30 Гц | Больше выражены симметрично с обеих сторон лобных областей головного мозга. Низкоамплитудные волны | - Состояние тревоги, пробуждения - Активная интенсивная умственная деятельность (мышление), тревожные мысли (размышления), активная концентрация (внимания) | Применение бензодиапинов |
| Гамма-ритм, γ | 30-100 Гц | Соматосенсорная кора больших полушарий головного мозга | - Появляется при сенсорном восприятии комбинации стимулов двух разных модальностей (например, свет и звук) - Проявляется при актуализации кратковременной памяти, необходимой для распознавания объектов (например, звук, тактильное воздействие | Уменьшение гамма-активности может быть связано со снижением познавательных возможностей |
| Мю-ритм, μ | 8-13 Гц | Сенсомоторная кора больших полушарий мозга | Проявляется в состоянии покоя мотонейронов | Подавление мю-ритма может указывать на активацию мотонейронов |
|
|
|
Таблица 13. Характеристика параметров электроэнцефалограммы и условия регистрации различных ритмов (Чеснокова, 2007)
| Наименование ритма | Частота, Гц | Аплитуда, мкВ | Условия регситрации ритма |
| Альфа-ритм | 8-13 | В состоянии умственного и физического покоя | |
| Бета-ритм | 13-30 | 20-25 | Эмоциональное возбуждение, умственная и физическая деятельность; при нанесении раздражений |
| Гамма-ритм | >30 | ||
| Тета-ритм | 4-8 | 100-150 | Сон, умеренные гипоксия и наркоз; при некоторых заболеваниях |
| Дельта ритм | 0,5-3,5 | 250-300 | Глубокий сон, наркоз и гипоксия; поражения коры больших полушарий |
Метод ВП — это регистрация колебаний электрической активности (электрического поля) мозга, возникающих на ЭЭГ при однократном раздражении сенсорных рецепторов (зрительных, слуховых, тактильных), афферентных путей, центров переключения афферентной импульсации, поступающей в кору большого мозга. Амплитуда их обычно невелика (≈15мкВ), сопоставима с волнами ЭЭГ, поэтому для эффективного выделения ВП применяют прием компьютерного суммирования и усреднения участков ЭЭГ (20—100 участков), следующих до и после включения раздражающего стимула. В процессе усреднения случайные колебания ЭЭГ трансформируются в изолинию, на фоне которой отчетливо проявляются закономерные колебания ВП (см. рис. 6.24). ВП регистрируют обычно при записи ЭЭГ. Поздние компоненты ВП обусловлены поступлением в кору неспецифических возбуждений через РФ ствола, неспецифические ядра таламуса и лимбической системы. Эту часть ВП называют вторичным ответом (см. рис. 6.24). Вторичные ответы, в отличие от первичных, регистрируются не только в первичных проекционных зонах, но и в других областях мозга, связанных между собой горизонтальными и вертикальными (кора — подкорка) нервными путями.
Метод ВП позволяет выявить взаимодействия различных зон коры при выработке условных рефлексов, исследовать ЭЭГ-корреляты в процессе психической деятельности.
Микроэлектродный метод позволяет изучать активность одиночных нейронов ЦНС. С помощью микроэлектродов, вводимых внутрь нервных клеток, можно измерять мембранные ПП, регистрировать постсинаптические потенциалы — возбуждающие и тормозные, а также ПД. Разновидностью микроэлектродного метода является метод микроионофореза, при котором используются многоканальные стеклянные микроэлектроды. Через один из каналов, заполненных электролитом, можно регистрировать электрическую активность нейрона; остальные каналы заполняют БАВ, которые апплицируют на работающий нейрон, пропуская через растворы веществ постоянный ток. Таким образом, в условиях прямого эксперимента с регистрацией активности одиночного нейрона можно наблюдать реакции последнего на действие различных агонистов и антагонистов, ДНК, РНК и их влияние на условные рефлексы и поведение животного, изучать механизмы памяти.
|
|
|
