 |


Механическая реакция целой мышцы при ее возбуждении
|
|
|
|
СРСП
По дисциплине: «Медицинская биофизика»
На тему:«Биомеханика мышц.Биофизика ммышечного сокращения»
Факультет: МПД
Группа: 103-Б
Выполнила: Шкунова М
Проверила: Оспанбекова Г.О
Алматы 2017
План
I Механизмы сокращения мышечного волокна.
II Источник энергии для сокращения мышечного волокна.
III Режимы и типы мышечных сокращений
Механизмы сокращения мышечного волокна
В покоящихся мышечных волокнах при отсутствии импульсации мотонейрона поперечные миозиновые мостики не прикреплены к актиновым миофиламентам. Тропомиозин расположен таким образом, что блокирует участки актина, способные взаимодействовать с поперечными мостиками миозина. Тропонин тормозит миозин — АТФ-азную активность и поэтому АТФ не расщепляется. Мышечные волокна находятся в расслабленном состоянии.
При сокращении мышцы длина А-дисков не меняется, J-диски укорачиваются, а Н-зона А-дисков может исчезать (рис. 4.3.).
 Рис.4.3. Сокращение мышцы. А — Поперечные мостики между актином и миозином разомкнуты. Мышца находится в расслабленном состоянии.
Рис.4.3. Сокращение мышцы. А — Поперечные мостики между актином и миозином разомкнуты. Мышца находится в расслабленном состоянии.
Б — Замыкание поперечных мостиков между актином и миозином. Совершение головками мостиков гребковых движений по направлению к центру саркомера. Скольжение актиновых нитей вдоль миозиновых, укорочение саркомера, развитие тяги.
Эти данные явились основой для создания теории, объясняющей сокращение мышцы механизмом скольжения (теорией скольжения) тонких актиновых миофиламентов вдоль толстых миозиновых. В результате этого миозиновые миофиламенты втягиваются между окружающими их актиновыми. Это приводит к укорочению каждого саркомера, а значит, и всего мышечного волокна.
|
|
|
Молекулярный механизм сокращения мышечного волокна состоит в том, что возникающий в области концевой пластинки потенциал действия распространяется по системе поперечных трубочек вглубь волокна, вызывает деполяризацию мембран цистерн саркоплазмати-ческого ретикулума и освобождение из них ионов кальция. Свободные ионы кальция в межфибриллярном пространстве запускают процесс сокращения. Совокупность процессов, обуславливающих распространение потенциала действия вглубь мышечного волокна, выход ионов кальция их саркоплазматического ретикулума, взаимодействие сократительных белков и укорочение мышечного волокна называют «электромеханическим сопряжением». Временная последовательность между возникновением потенциала действия мышечного волокна, поступлением ионов кальция к миофибриллам и развитием сокращения волокна показана на рисунке 4.4.
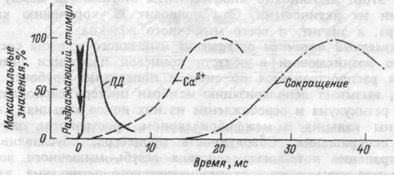 Рис.4.4. Схема временной последовательности развития
Рис.4.4. Схема временной последовательности развития
потенциала действия (ПД), освобождения ионов кальция (Са2+) и развития изометрического сокращения мышцы.
При концентрации ионов Са2+ в межмиофибриллярном пространстве ниже 10″ тропомиозин располагается таким образом, что блокирует прикрепление поперечных миозиновых мостиков к нитям актина. Поперечные мостики миозина не взаимодействуют с нитями актина. Продвижение относительно друг друга нитей актина и миозина отсутствует. Поэтому мышечное волокно находится в расслабленном состоянии. При возбуждении волокна Са2+выходит из цистерн саркоплазматического ретикулума и, следовательно, концентрация его вблизи миофибрилл возрастает. Под влиянием активирующих ионов Са2+ молекула тропонина изменяет свою форму таким образом, что выталкивает тропомиозин в желобок между двумя нитями актина, освобождая тем самым участки для прикрепления миозиновых поперечных мостиков к актину. В результате поперечные мостики прикрепляются к актиновым нитям. Поскольку головки миозина совершают «гребковые» движения в сторону центра саркомера происходит «втягивание» актиновых миофиламентов в промежутки между толстыми миозиновыми нитями и укорочение мышцы.
|
|
|
Источник энергии для сокращения мышечных волокон
Источником энергии для сокращения мышечных волокон служит АТФ. С инактивацией тропонина ионами кальция активируются каталитические центры для расщепления АТФ на головках миозина. Фермент миозиновая АТФ-аза гидролизует АТФ, расположенный на головке миозина, что обеспечивает энергией поперечные мостики. Освобождающиеся при гидролизе АТФ молекула АДФ и неорганический фосфат используются для последующего ресинтеза АТФ. На миозиновом поперечном мостике образуется новая молекула АТФ. При этом происходит разъединение поперечного мостика с нитью актина. Повторное прикрепление и отсоединение мостиков продолжается до тех пор, пока концентрация кальция внутри миофибрилл не снижается до подпороговой величины. Тогда мышечные волокна начинают расслабляться.
При однократном движении поперечных мостиков вдоль актиновых нитей (гребковых движениях) саркомер укорачивается примерно на 1% его длины. Следовательно, для полного изотонического сокращения мышцы необходимо совершить около 50 таких гребковых движений. Только ритмическое прикрепление и отсоединение головок миозина может втянуть нити актина вдоль миозиновых и совершить требуемое укорочение целой мышцы. Напряжение, развиваемое мышечным волокном, зависит от числа одновременно замкнутых поперечных мостиков. Скорость развития напряжения или укорочения волокна определяется частотой замыкания поперечных мостиков, образуемых в единицу времени, то есть скоростью их прикрепления к актиновым миофиламентам. С увеличением скорости укорочения мышцы число одновременно прикрепленных поперечных мостиков в каждый момент времени уменьшается. Этим и можно объяснить уменьшение силы сокращения мышцы с увеличением скорости ее укорочения.
При одиночном сокращении процесс укорочения мышечного волокна заканчивается через 15-50 мс, так как активирующие его ионы кальция возвращаются при помощи кальциевого насоса в цистерны саркоплазматического ретикулума. Происходит расслабление мышцы.
|
|
|
Поскольку возврат ионов кальция в цистерны саркоплазматического ретикулума идет против диффузионного градиента, то этот процесс требует затрат энергии. Ее источником служит АТФ. Одна молекула АТФ затрачивается на возврат 2-х ионов кальция из межфибриллярного пространства в цистерны. При снижении содержания ионов кальция до подпорогового уровня (ниже 10 V) молекулы тропонина принимают форму, характерную для состояния покоя. При этом вновь тропомиозин блокирует участки для прикрепления поперечных мостиков к нитям актина. Все это приводит к расслаблению мышцы вплоть до момента прихода очередного потока нервных импульсов, когда описанный выше процесс повторяется. Таким образом, кальций в мышечных волокнах играет роль внутриклеточного посредника, связывающего процессы возбуждения и сокращения.
Режимы и типы мышечных сокращений
Одиночное сокращени
Режим сокращений мышечных волокон определяется частотой импульсации мотонейронов. Механический ответ мышечного волокна или отдельной мышцы на однократное их раздражение называется одиночным сокращением.
При одиночном сокращении выделяют:
1. Фазу развития напряжения или укорочения;
2. Фазу расслабления или удлинения (рис.4.5.).
 Рис.4.5. Развитие во времени потенциала действия (А) и изометрического сокращения мышцы, приводящей большой палец кисти (Б).
Рис.4.5. Развитие во времени потенциала действия (А) и изометрического сокращения мышцы, приводящей большой палец кисти (Б).
1 — фаза развития напряжения; 2 — фаза расслабления.
Фаза расслабления продолжается примерно в два раза дольше, чем фаза напряжения. Длительность этих фаз зависит от морфофункциональных свойств мышечного волокна: у наиболее быстро сокращающихся волокон глазных мышц фаза напряжения составляет 7-10 мс, а у наиболее медленных волокон камбаловидной мышцы — 50-100 мс.
В естественных условиях мышечные волокна двигательной единицы и скелетная мышца в целом работают в режиме одиночного сокращения только в том случае, когда длительность интервала между последовательными импульсами мотонейрона равна или превышает длительность одиночного сокращения иннервируемых им мышечных волокон. Так, режим одиночного сокращения медленных волокон камбаловидной мышцы человека обеспечивается при частоте импульсации мотонейрона менее 10 имп/с, а быстрых волокон глазодвигательных мышц — при частоте импульсации мотонейрона менее 50 имп/с.
|
|
|
В режиме одиночного сокращения мышца способна работать длительное время без развития утомления. Однако в связи с тем, что длительность одиночного сокращения невелика, развиваемое мышечными волокнами напряжение не достигает максимально возможных величин. При относительно высокой частоте импульсации мотонейронов каждый последующий раздражающий импульс приходится на фазу предшествующего напряжения волокона, то есть до того момента, когда оно начинает расслабляться. В этом случае механические эффекты каждого предыдущего сокращения суммируются с последующим. Причем величина механического ответа на каждый последующий импульс меньше, чем на предыдущий. После нескольких первых импульсов последующие ответы мышечных волокон не изменяют достигнутого напряжения, а лишь поддерживают его. Такой режим сокращения называется гладким тетанусом (рис.4.6.). В подобном режиме двигательные единицы мышц человека работают при развитии максимальных изометрических усилий. При гладком тетанусе развиваемое ДЕ напряжение в 2-4 раза больше, чем при одиночных сокращениях.
 Рис.4.6. Одиночные (а) и тетанические (б,в,г,д) сокращения скелетной мышцы. Накладывание волн сокращения друг на друга и образование тетануса при частотах раздражения: 5 -15 раз/с; в — 20 раз/с; г — 25 раз/с; д — более 40 раз в 1 сек (гладкий тетанус).
Рис.4.6. Одиночные (а) и тетанические (б,в,г,д) сокращения скелетной мышцы. Накладывание волн сокращения друг на друга и образование тетануса при частотах раздражения: 5 -15 раз/с; в — 20 раз/с; г — 25 раз/с; д — более 40 раз в 1 сек (гладкий тетанус).
В тех случаях, когда промежутки между последовательными импульсами мотонейрона меньше времени полного цикла одиночного сокращения, но больше длительности фазы напряжения, сила сокращения ДЕ колеблется. Этот режим сокращения называется зубчатым тетанусом (рис. 4.6.).
Гладкий тетанус для быстрых и медленных мыши достигается при разных частотах импульсации мотонейронов. Зависит это от времени одиночного сокращения. Так, гладкий тетанус для быстрой глазодвигательной мышцы проявляется при частотах свыше 150-200 имп/с, а у медленной камбаловидной мышцы — при частоте около 30 имп/с. В режиме тетанического сокращения мышца способна работать лишь короткое время. Это объясняется тем, что из-за отсутствия периода расслабления она не может восстановить свой энергетический потенциал и работает как бы «в долг».
Механическая реакция целой мышцы при ее возбуждении
|
|
|
Механическая реакция целой мышцы при ее возбуждении выражается в двух формах — в развитии напряжения и в укорочении. В естественных условиях деятельности в организме человека степень укорочения мышцы может быть различной.
По величине укорочения различают три типа мышечного сокращения:
1. Изотонический — это сокращение мышцы, при которой ее волокна укорачиваются при постоянной внешней нагрузке. В реальных движениях чисто изотоническое сокращение практически отсутствует;
2. Изометрический — это тип активации мышцы, при котором она развивает напряжение без изменения своей длины. Изометрическое сокращение лежит в основе статической работы;
3. Ауксотонический или анизотонический тип — это режим, в котором мышца развивает напряжение и укорачивается. Именно такие сокращения имеют место в организме при естественных локомоциях — ходьбе, беге и т.д.
Динамическое сокращени
Изотонический и анизотонический типы сокращения лежат в основе динамической работы локомоторного аппарата человека.
При динамической работе выделяют:
1. Концентрический тип сокращения — когда внешняя нагрузка меньше, чем развиваемое мышцей напряжение. При этом она укорачивается и вызывает движение;
2. Эксцентрический тип сокращения — когда внешняя нагрузка больше, чем напряжение мышцы. В этих условиях мышца, напрягаясь, все же растягивается (удлиняется), совершая при этом отрицательную (уступающую) динамическую работу
|
|
|
