 |


Содержание. ВВЕДЕНИЕ. 3. Механизм действия гормонов
|
|
|
|
Министерство сельского хозяйства и продовольствия Республики Беларусь
Витебская ордена «Знак Почета» государственная академия ветеринарной медицины
Кафедра химии
«БИОХИМИЯ ГОРМОНОВ»
(УЧЕБНО - МЕТОДИЧЕСКОЕ ПОСОБИЕ)
ВИТЕБСК 2002
Авторы: Котович И. В., старший преподаватель
Германович Н. Ю., кандидат биологических наук, доцент
Рецензенты: Матузко Н. С., кандидат биологических наук, доцент
Шевченко И. С., кандидат ветеринарных наук, доцент
Биохимия гормонов (учебно-методическое пособие)
Рассмотрено и рекомендовано к печати учебно-методической комиссией факультета ветеринарной медицины 9. 04. 2002 г., протокол № 2
Рекомендовано к печати редакционно-издательским Советом академии
Содержание
Стр.
| Введение | |
| 1. Общее представление о гормонах | |
| 2. Классификация гормонов | |
| 3. Механизм действия | |
| 4. Строение и биологическая роль гормонов: | |
| 4. 1. гипофиза | |
| 4. 2. гипоталамуса | |
| 4. 3. щитовидной железы | |
| 4. 4. паращитовидных желез | |
| 4. 5. поджелудочной железы | |
| 4. 6. надпочечников | |
| 4. 7. половых желез | |
| 4. 8. плаценты | |
| 4. 9 тимуса | |
| 4. 10. эпифиза | |
| 5. Биологически активные вещества гормоноподобного действия | |
| Литература |
ВВЕДЕНИЕ
|
|
|
Одной из удивительных особенностей живых организмов является их способность сохранять постоянство внутренней среды – гомеостаз – при помощи механизмов саморегуляции, в которых одно из главных мест принадлежит гормонам. У высших животных координированное протекание всех биологических процессов не только в целостном организме, но и в микропространстве отдельной клетки и даже в отдельном субклеточном образовании (митохондрии, микросомы) определяется нейрогуморальными механизмами, сложившимися в процессе эволюции. При помощи этих сигналов организм воспринимает разнообразные сигналы об изменениях в окружающей и внутренней средах и тонко регулирует интенсивность процессов обмена. В регуляции этих процессов, в осуществлении протекания множества реакций гормоны занимают промежуточное звено между нервной системой и действием ферментов, которые непосредственно регулируют скорость обмена веществ. Любые нарушения синтеза или распада гормонов, вызванные разнообразными причинными факторами, включая заболевания эндокринных желез (состояние гипо- и гиперфункции) или изменения структуры и функций рецепторов и внутриклеточных посредников, приводят к изменению нормального синтеза ферментов и соответственно к нарушению метаболизма.
Учение о гормонах выделено в самостоятельную науку – эндокринологию. Современная эндокринология изучает химическую структуру гормонов, образующихся в железах внутренней секреции, зависимость между структурой и функцией гормонов, молекулярные механизмы действия, а также физиологию и патологию эндокринной системы. В наши дни эндокринология превратилась в один из самых бурно развивающихся разделов биологической науки. Она имеет свои цели и задачи, специфические методологические подходы и методы исследования.
Настоящее учебно-методическое пособие предназначено для студентов факультета ветеринарной медицины и зооинженерного факультета. Оно содержит сведения о номенклатуре и классификации гормонов, их химической природе, биологической роли. Раздел «механизм действия гормонов» проиллюстрирован схемами, что позволяет лучше осмыслить изучаемый материал. В пособии также рассматриваются нарушения обменных процессов, возникающих при гипо- и гиперфункциях эндокринных желез.
|
|
|
Материал пособия дает возможность студентам приобрести знания, необходимые для изучения закономерностей протекания метаболических процессов в организме, что позволит понять патогенез многих заболеваний и влияние гормонов на продуктивность животных.
1. Общее представление о гормонах
Характерная особенность многоклеточных организмов – наличие дифференцированных тканей. Последние обладают специализированными функциями, обеспечивающими выживание организма.
Для координации тканевых ответов на изменение условий внешней и внутренней среды необходимы механизмы межклеточной коммуникации. В ходе эволюции сформировались две основные системы, выполняющие эту функцию: нервная, для которой характерны высокая скорость распространения сигнала, строгая направленность и кратковременность действия и эндокринная, для которой характерны наличие разнообразных гормонов, которые секретируются специфическими железами и затем транспортируются, воздействуя на прилежащие или удаленные ткани. Эти две системы перекрываются и накладываются друг на друга. Важную роль играет нервная регуляция эндокринной функции; многие нейромедиаторы (катехоламины, ацетилхолин) сходны с гормонами по способу синтеза, высвобождения, транспорту и механизму действия.
Гормоны(от греч. hormaino – побуждаю) - это биологически активные вещества, синтезируемые специализированными клетками и железами внутренней секреции, поставляемые кровью в другие органы и ткани, где оказывают регулирующее влияние на обмен веществ и физиологические функции.
Однако исходное определение слишком узко. Установлено, что гормоны способны воздействовать на прилегающие клетки данной ткани (паракринный эффект), а также на клетки, в которых они синтезируются (аутокринный эффект).
|
|
|
Учение о гормонах выделено в самостоятельную науку - эндокринологию (изучает химическую структуру гормонов, зависимость между структурой и функцией, механизм действия, физиологию и патологию эндокринной системы).
Гормонам присущи следующие общие биологические признаки.
1. Специфичность - своеобразие его химической структуры, функции и места образования; Подобные вещества выделяют также клетки некоторых органов, не относящихся к железам внутренней секреции (клетки желудочно-кишечного тракта, тучные клетки соединительной ткани, клетки почек, клетки эндотелия и др. ). В отличие от гормонов эти биологически активные вещества действуют в местах образования и обозначаются как гормоноиды (гормоноподобные вещества).
2. Высокая биологическая активность (эффект проявляется в присутствии чрезвычайно малых концентраций гормона (10-11 – 10-6 М).
3. Секретируемость – ряд специфических, биологически активных веществ может образовываться в эндокринной железе в качестве промежуточных продуктов биосинтеза гормонов. Например, 11-дезоксикортизол, образующийся в коре надпочечников и обладающий биологической активностью, в норме не секретируется в кровь.
4. Дистанционность действия, т. е. гормоны регулируют обмен веществ и функции эффекторных клеток на расстоянии.
В основе каждой эндокринной функции (кортикостероидной, тиреоидной, соматотропной и т. д. ) лежит образование специфических гормонов в соответствующей железе. Однако, биосинтез и секреция гормонов не исчерпывают понятия «эндокринная функция». Эндокринная функция – сложноорганизованная система, состоящая из следующих компонентов:
- биосинтез и секреция гормонов (или прогормонов) в железе;
- процессы специфической регуляции и саморегуляции функций железы;
- специфический транспорт секретируемых гормонов в крови;
- система специфического метаболизма гормонов в периферических тканях и их экскреция;
- специфическое взаимодействие гормонов с реагирующими тканями.
Биосинтез и секреция гормонов. Центральным звеном любой эндокринной функции является продукция гормонов соответствующей железой. Продукция гормонов железой складывается из двух процессов.
|
|
|
1. Биосинтез гормона, в результате которого формируется его химическая структура.
2. Секреция образовавшегося гормона из железы в кровь.
Биосинтез гормонов - это цепь биохимических реакций, приводящих к формированию специфической структуры гормональной молекулы. Как химическая природа активных гормонов, так и механизмы их биосинтеза весьма разнообразны – могут образовываться из липидных предшественников, в результате модификации аминокислоты тирозина, путем белкового синтеза. Некоторые гормоны синтезируются в своей конечной форме (альдостерон, гидрокортизон, эстрадиол, катехоламины), другие перед секрецией или для приобретения полной биологической активности должны подвергаться модификации (инсулин синтезируется в виде проинсулина, у паратгормона есть по крайней мере два пептида-предшественника, содержащие последовательности, отщепление которых необходимо для проявления полной биологической активности).
Генетический контроль биосинтеза осуществляется в зависимости от природы синтезируемого гормона:
- либо на уровне иРНК самого гормона (в случае белково-пептидных гормонов);
- либо на уровне ферментов, которые осуществляют биосинтез того либо иного гормона (для гормональных стероидов, производных аминокислот и небольших пептидов типа рилизинг-факторов).
Секреция гормонов представлена совокупностью процессов, обусловливающих освобождение синтезированных гормональных соединений из эндокринных клеток в венозную кровь и лимфу. Секреторные процессы протекают спонтанно, обеспечивая некий базальный уровень гормонов в циркулирующих жидкостях. Секреторный процесс осуществляется железистыми клетками отдельными дискретными порциями. “Квантовый“ характер гормональной секреции обусловлен, по-видимому, циклическим характером процессов биосинтеза, внутриклеточного депонирования и траспорта гормонов. По особенностям механизмов секреторные процессы можно разделить на 3 типа.
1. Освобождение гормонов из клеточных секреторных гранул (секреция белково-пептидных гормонов и катехоламинов).
2. Освобождение гормонов из белковосвязанной формы (секреция тиреоидных гормонов).
3. Относительно свободная диффузия гормонов через клеточные мембраны (стероидные гормоны).
Катаболизм гормонов. Катаболизм гормонов - это совокупность процессов ферментативной деградации исходной химической структуры гормональных соединений. В катаболизирующих органах наряду с необратимой инактивацией могут протекать обменные процессы, приводящие к активации, реактивации, взаимопревращениям гормонов и возникновению новой гормональной активности. К активации относится превращение тироксина в трииодтиронин, к реактивации - переход кортизона в кортизол, примером взаимопревращений гормонов разного типа являются превращения андрогенов в эстрогены в гипоталамусе. В качестве показателя интенсивности метаболических процессов на уровне целого организма используется величина периода полураспада. Это время, за которое концентрация введенной в кровь порции радиоактивного гормона необратимо уменьшается вдвое. Тиреоидные гормоны, в особенности тироксин, катаболизируется очень медленно (сутки), а катехоламины - исключительно быстро (секунды, минуты). Основные пути катаболизма для стероидных гормонов – это восстановление двойной связи в кольце А, окисление-восстановление некоторых кислородсодержащих функциональных групп и гидроксилирование углеродных атомов. Для катехоламинов существует два доминирующих пути – это окислительной дезаминирование боковой цепи и оксиметилирование гидроксила у С3 атома кольца. Периферический метаболизм белково-пептидных гормонов изучен в настоящее время недостаточно. Однако, известно, что они быстро исчезают из крови вследствие работы специфических пептидаз. В составе выводимых жидкостей плохо растворимые в воде метаболиты стероидных гормонов экскретируются в основном в форме глюкуронидов, сульфатов и некоторых других эфиров, обладающих высокой водорастворимостью. Метаболиты аминокислотных гормонов экскретируются главным образом в свободной форме. Метаболиты белково-пептидных гормонов выводятся преимущественно в форме свободных аминокислот или их солей и небольших пептидов. Гормональные метаболиты экскретируются в первую очередь с мочой и желчью.
|
|
|
2. Классификация гормонов
По химической природе гормоны можно разделить на 3 группы:
a) гормоны белковой и пептидной природы (гормоны гипофиза, поджелудочной железы, паращитовидных желез, тимуса);
b) низкомолекулярные производные аминокислот (гормоны мозгового слоя надпочечников и иодтиронины щитовидной железы);
c) гормоны стероидной природы (гормоны половых желез и коркового слоя надпочечников).
3. Механизм действия гормонов
В управлении метаболизмом гормоны участвуют следующим образом. Поток информации о состоянии внутренней среды организма и об изменениях, связанных с внешними воздействиями поступает в нервную систему, там перерабатывается и формируется ответный сигнал. Он поступает к органам–эффекторам в виде нервных импульсов по цетробежным нервам и опосредованно через эндокринную систему.
Пунктом, где сливаются потоки нервной и эндокринной информации является гипоталамус – сюда поступают нервные импульсы из разных отделов головного мозга. Они определяют продукцию и секрецию гипоталамических гормонов, влияющих в свою очередь через гипофиз на продукцию гормонов периферическими эндокринными железами. Гормоны периферических желез, в частности мозгового вещества надпочечников, контролируют секрецию гипоталамических. В конечном счете, содержание гормона в кровотоке поддерживается по принципу саморегуляции. Высокий уровень гормона выключает или ослабляет по механизму отрицательной обратной связи его образование, низкий уровень усиливает продукцию.
Гормоны действуют на ткани избирательно, что обусловлено неодинаковой чувствительностью к ним тканей. Органы и клетки, наиболее чувствительные к влиянию определенного гормона, принято называть мишенью гормона (орган-мишень или клетка-мишень).
Концепция ткани- мишени. Ткань-мишень – это такая ткань, в которой гормон вызывает специфическую физиологическую (биохимическую) реакцию Общую реакцию ткани- мишени на действие гормона определяет целый ряд факторов. Прежде всего это локальная концентрация гормона вблизи ткани-мишени, зависящая от:
1. скорости синтеза и секреции гормона;
2. анатомической близости ткани-мишени к источнику гормона;
3. констант связывания гормона со специфическим белком-переносчиком (если такой существует);
4. скорости превращения неактивной или малоактивной формы гормона в активную;
5. скорости исчезновения гормона из крови в результате распада или выведения.
Собственно тканевой ответ определяется:
- относительной активностью и (или) степенью занятости специфических рецепторов
- состоянием сенситизации - десентизации клетки.
Специфичность гормонов по отношению к клеткам-мишеням обусловлена нర личием у клеток специфических р䐵 цепторов.
Все рецепторы гормонов можно разделить на 2 䑂 ипа:
1) локализованныеࠠ на наружной поверхности клеточной мембраны;
2) расположенные в цитоплазме клетки.
Свойства рецепторов:
- четкая субстратная специфичность;
- насыщаемость;
- сродство к гормону в пределе биологических концентраций гормона;
- обратимость действия.
В зависимости от того, где в клетке происходит передача информации, можно выделить следующие варианты действия гормонов:
1) Мембранный (локальный).
2) Мембранно-внутриклеточный или опосредованный.
3) Цитоплазматический (прямой).
Мембранный тип действия реализуется в месте связывания гормона с плазматической мембраной и заключается в избирательном изменении ее проницаемости. По механизму действия гормон в данном случае выступает как аллостерический эффектор транспортных систем мембраны. Так, например, обеспечивается трансмембранный перенос глюкозы под действием инсулина, аминокислот и некоторых ионов. Обычно мембранный тип действия сочетается с мембранно-внутриклеточным.
Мембранно-внутриклеточное действие гормонов характеризуется тем, что гормон не проникает в клетку, а влияет на обмен в ней через посредник, который является как бы представителем гормона в клетке – вторичным посредником (первичный посредник – сам гормон). По типу вторичных посредником действуют циклические нуклеотиды (цАМФ, цГМФ) и ионы кальция.
 |
Регуляция – сложный комплексный механизм, реагирующий на различного рода воздействия изменением обмена веществ и поддерживающий постоянство внутренней среды.
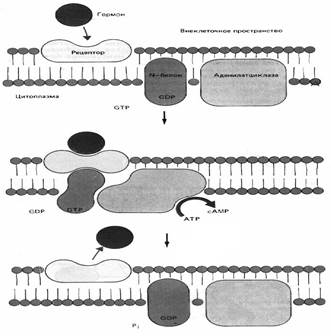
Регуляция через цАМФ или цГМФ. В цитоплазматическую мембрану клетки встроен фермент аденилатциклаза, состоящий из 3-х частей – узнающей (набор рецепторов, располагающихсяна поверхности мембраны), сопрягающей (N-белок, занимающий в липидном бислое мембраны промежуточное положение между рецептором и каталитической частью) и каталитический (собственно ферментный белок, активный центр которого обращен внутрь клетки). В каталитическом белке имеются раздельные участки для связывания цАМФ и цГМФ.
Передача информации, источником которой является гормон, происходит следующим образом:
- гормон связывается с рецептором;
- комплекс гормон-рецептор взаимодействует с N-белком, изменяя его конфигурацию;
- изменение конфигурации приводит к превращению ГДФ (присутствующим в неактивном белке) в ГТФ;
- комплекс белок-ГТФ активирует собственно аденилатциклазу;
- активная аденилатциклаза нарабатывает цАМФ внутри клетки (АТФ ¾ ® цАМФ + H4P2O7)
Аденилатциклаза работает до тех пор, пока сохраняется комплекс гормон-рецептор, поэтому одна молекула комплекса успевает образовывать от 10 до 100 молекул цАМФ.
Синтез цГМФ запускается таким же путем, с той лишь разницей, что комплекс гормон-рецептор активирует гуанилатциклазу, продуцирующую цГМФ из ГТФ.
Далее процессы происходят следующие процессы:
- циклические нуклеотиды активируют протеинкиназы (цАМФ-зависимые или цГМФ-зависимые);
- активированные протеинкиназы фосфорилируют за счет АТФ разные белки;
- фосфорилирование сопровождается изменением функциональной активности (активацией или угнетением) этих белков.
Циклические нуклеотиды (цАМФ и цГМФ) действуют на разные белки, поэтому эффект зависит от мембранного рецептора, связывающего гормон. Характер рецептора определяет, будет буде изменена активность цАМФ- или цГМФ-зависимых белков-ферментов. Нередко эти нуклеотиды оказывают противоположные эффекты. Поэтому биохимические процессы в клетке под влиянием одного гормона могут активироваться или угнетаться в зависимости от того, какими рецепторами располагает клетка. Например, адреналин может связываться b- и a-рецепторами. Первые включают аденилатциклазу и образование цАМФ, вторые – гуанилатциклазу и образование ц ГМФ. Циклические нуклеотиды активируют разные белки, поэтому характер метаболических изменений в клетке зависит не от гормона, а от рецепторов, которыми располагает клетка.
Влияние циклических нуклеотидов на метаболизм прекращается с помощью ферментов фосфодиэстераз.
Таким образом, процесс управляемый через аденилатциклазную систему, зависит от соотношения между скоростью продукции цАМФ или цГМФ и скоростью их распада.
Механизм действия гормонов, включающий аденилатциклазную систему, присущ гормонам белковой и полипептидной природы, а также катехоламинам (адреналину, норадреналину).
Цитоплазматический механизм действия присущ гормонам стероидной природы.
Рецепторы стероидных гормонов расположены в цитоплазме клетки. Эти гормоны (обладающие липофильными свойствами), проникая в клетку, взаимодействуют с рецепторами с образованием гормон-рецепторного комплекса, который после молекулярной перестройки, приводящей к его активации, поступает в ядро клетки, где взаимодействует с хроматином. При этом происходит активация генов и в последующем развивается цепь процессов, сопровождающихся усиленным синтезом РНК, в том числе информационных. Это приводит к индукции соответствующих ферментов в ходе процесса трансляции, что влечет за собой изменение скорости и направленности метаболических процессов в клетке.
Таким образом, в этом случае гормональный эффект реализуется на уровне генетического аппарата клетки-мишени.
Биологические эффекты гормонов, воздействующих на генетический аппарат клетки, проявляются главным образом во влиянии на рост и дифференцировку тканей и органов.
Смешанный тип передачи информации свойственен иодтиронинам (гормонам щитовидной железы), которые по липофильным свойствам занимают промежуточное положение между водорастворимыми и липофильными (стероидными) гормонами. Эта группа гормонов реализует свой эффект и мембранно-внутриклеточным и цитозольным механизмами.
|
|
|
