 |


Генетическая характеристика прокариот и эукариот
|
|
|
|
Министерство здравоохранения и социального развития РФ
Государственное бюджетное образовательное учреждение
Высшего профессионального образования
Читинская государственная медицинская академия
УТВЕРЖДАЮ
Зав. кафедрой ________________Клеусова Н.А.
ТЕМА: Механизм реализациии наследственной информации в признак
методические указания для студентов
лечебного факультета
Составила к.б.н., ассистент Чистякова Н.С.
Чита-2014
Тема: Механизм реализациии наследственной информации в признак
Цель: при изучении данной темы формируются компетенции ОК-1, ПК-31, ПК-32, изучив данную тему, студент должен
Знать
· строение, свойства, функции ДНК
· виды РНК, строение, функции
· реакции матричного синтеза (транскрипция, трансляция)
Уметь
· моделировать процесс кодирования наследственной информации
· моделировать этапы реализации наследственной информации
Владеть
· методикой решения задач
Задание для самоподготовки
1. Раскрыть содержание «Центральной догмы молекулярной биологии».
2. Химический состав, структуры и свойства молекулы ДНК.
3. Химический состав, структуры молекулы РНК, охарактеризовать виды РНК клетки.
4. Охарактеризовать первичную, вторичную, третичную и четвертичную структуру белковой молекулы.
5. Генетический код, единицы кода, свойства кода.
6. Матричный синтез белка и его этапы: транскрипция, трансляция.
7. Знать термины: кодон, антикодон, коллинеарность, процессинг, матрица.
Содержание темы
Нуклеиновые кислоты
Материальным субстратом наследственности является ДНК. ДНК – высокомолекулярное полимерное соединение, мономерами которого являются нуклеотиды, которые включают три компонента (рис. 1):
|
|
|
1) пятиуглеродный сахар – дезоксирибоза
2) остаток фосфорной кислоты
3) одно из четырех азотистых оснований (аденин, тимин, гуанин, цитозин).

Рис. 1. Состав нуклеотида
Нуклеотиды соединяются в полинуклеотидную цепь с помощью фосфо-диэфирных связей. Сборка осуществляется с помощью фермента полимеразы. Каждый последующий нуклеотид присоединяется к гидроксильной группе стоящей в положении атома углерода 3´. Начало цепи несет фосфатную группу в положении 5´. Это позволяет в полинуклеотидной цепи выделить 5´ и 3´концы.
В структурной организации молекулы ДНК можно выделить:
1) первичную структуру –.одинарную полинуклеотидную цепь (рис. 2);
2) в торичную структуру - две комплементарные друг другу и антипараллельные полинуклеотидные цепи, соединенные водородными связями;
3) третичную структуру – предложили в 1953 г. американский биофизик Дж. Уотсон и английский биофизик Ф. Крик (рис. 3).
Данные рентгеноструктурного анализа показали, что молекула ДНК состоящая из двух цепей, образует спираль, закрученную вокруг собственной оси. Диаметр спирали составляет 2 нм, длина витка 3,4 нм. В каждый виток входит 10 пар нуклеотидов. Двойные спирали ДНК могут быть право и левозакрученными. Большинство молекул ДНК находятся в правозакрученной В-форме (В-ДНК).

Рис. 2. Первичная структура ДНК
Однако встречаются и левозакрученные формы (Z-ДНК). Значение этой формы пока не установлено (рис. 3)
Одним из важнейших свойств ДНК является способность к репликации. В процессе репликации каждая из двух цепей материнской молекулы служит матрицей для дочерней. После репликации вновь синтезированная молекула ДНК содержит одну «материнскую» цепочку, а вторую «дочернюю» - вновь синтезированную. Такой способ самоудвоения (рис.4) называется полуконсервативным.
|
|
|

Рис. 3. Пространственные модели левозакрученной Z-формы (I) и правозакрученной В-формы (II) ДНК

Рис. 4. Принцип полуконсервативной репликации ДНК.
Для матричного синтеза новой молекулы ДНК необходимо, чтобы старая молекула была деспирализована и вытянута. С помощью фермента геликазы двойная спираль ДНК в отдельных зонах (репликонах) расплетается. Области расхождения полинуклеотидных цепей в зонах репликации называют (рис. 5) репликационными вилками.
В каждой такой области с помощью фермента ДНК – полимеразы синтезируется ДНК двух новых дочерних молекул. В каждом репликоне ДНК-полимераза может двигаться вдоль материнской нити только в одном направлении (5´ - 3´), поэтому процесс репликации при антипараллельном соединении двух цепей ДНК будет протекать по-разному. На одной из матриц 3´ - 5´ сборка новой цепи происходит непрерывно от 5´к 3´концу и она постепенно удлиняется на 3´ - конце то другая цепь, синтезируемая на матрице 5´ - 3´конце должна была бы расти от 3´ к 5´ концу, но это противоречит направлению действия фермента ДНК-полимеразы. В настоящее время установлено, что синтез второй цепи ДНК осуществляется короткими фрагментами также в направлении от 5´к 3´ концу (рис. 6) (фрагменты Оказаки). Фрагменты Оказаки сшиваются в единую нить ферментом лигазой. Из двух синтезируемых дочерних цепей одна строится непрерывно, ее синтез идет быстрее - эту цепь называют лидирующей. Синтез другой идет медленнее.

Рис. 5. Образование репликационной вилки ДНК

Рис. 6. Синтез лидирующей и отстающей цепи.
Рибонуклеиновая кислота (РНК). Нуклеотид РНК состоит из одного из четырех азотистых аснований (аденина, гуанина, цитозина или урацила), углевода рибозы и остатка фосфорной кислоты. Молекулы РНК одноцепочные.
Виды РНК. Рибосомальная РНК (р-РНК) в соединении с белком входит в состав рибосом. Информационная РНК (и-РНК) синтезируется на полуцепи ДНК, несет информацию о синтезе определенного белка. Она представляет собой одиночную полинуклеотидную цепь. Мономерами РНК являются также 4 нуклеотида – адениловый, гуаниловый, вместо тимилового – урациловый и цитозиновый. Вместо углевода дезоксирибозы в состав нуклеотидов РНК входит углевод рибоза. Транспортная РНК (т-РНК), имеет форму трилистника (рис. 7), имеет короткую нуклеотидную последовательность. В ней выделяют четыре главные части: акцепторный стебель - одноцепочный участок, который заканчивается последовательностью нуклеотидов ЦЦА со свободной ОН группой. К этому концу присоединяется транспортируемая аминокислота, боковые петли, средняя петля антикодоновая – состоит из 5-ти нуклеотидов и содержит в центре антикодон. Антикодон - это три нуклеотида, который шифруют аминокислоту, транспортируемую к месту синтеза белка. Каждая тРНК может переносить только свою аминокислоту. Процесс узнавания своей аминокислоты тРНК называется рекогницией. Аминокислота присоединяется к аденину триплета ЦЦА с помощью фермента аминоацил-тРНК-синтетазы. Образуется комплекс аминоацил тРНК. Служит для транспорта аминокислот к месту синтеза белка.
|
|
|

Рис. 7. Строение тРНК
Информация о первичной структуре белковой молекулы закодирована в ДНК в виде генетического кода (табл. 2). Система расположения нуклеотидов в цепи ДНК, контролирующая последовательность расположения аминокислот в белковой цепи называется генетическим кодом. Расшифровка генетического кода была начата в 1954 г. Г. Гамовым.
Свойства генетического кода:
1. Триплетность - одной аминокислоте в полипептидной цепочке соответствуют три рядом расположенных нуклеотида; минимальная единица функции – триплет (кодон).
2. Вырожденность (избыточность) – количество возможных триплетов 64 (61 кодирующие, и 3 нонсенс – в ДНК АТТ, АЦТ, АТЦ, в иРНК УАА, УГА, УАГ), а аминокислот 20, поэтому одну аминокислоту может кодировать несколько триплетов (табл. 1)
3. Специфичность – каждый триплет кодирует только одну аминокислоту
4. Неперекрываемость -один нуклеотид входит в составтолько одного триплета
5. Универсальность – у всех живых организмов одинаковые триплеты кодируют одинаковые аминокислоты
Наследственная информация записанная с помощью генетического кода хранится в ДНК и размножается для того, чтобы обеспечить новые клетки генетическим материалом, который является «инструкцией» для их нормального развития и функционирования. Непосредственного участия ДНК в синтезе белка не принимает. Роль посредника выполняет иРНК.
|
|
|
Таблица 1
Генетический код
| У | Ц | А | Г | ||
| У | Фен Фен Лей Лей | Сер Сер Сер Сер | Тир Тир Non 2 Non 1 | Цис Цис Non 3 Три | У Ц А Г |
| Ц | Лей Лей Лей Лей | Про Про Про Про | Гис Гис Глн Глн | Арг Арг Арг Арг | У Ц А Г |
| А | Иле Иле Иле Мет | Тре Тре Тре Тре | Асн Асн Лиз Лиз | Сер Сер Арг Арг | У Ц А Г |
| Г | Вал Вал Вал Вал | Ала Ала Ала Ала | Асп Асп Глу Глу | Гли Гли Гли Гли | У Ц А Г |
Биосинтез белка у прокариот идет по схеме:
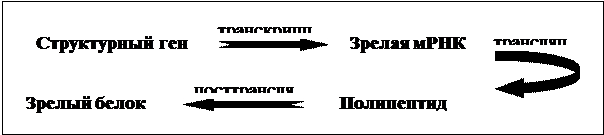
У эукариот процесс синтеза белка пойдет иначе:

Транскрипция. Для того чтобы синтезировался белок с заданными свойствами, происходит переписывание информации с ДНК на иРНК. Этот этап называется транскрипцией.Транскрипция подразделяется на три основных стадии – инициацию (начало синтеза мРНК), элонгацию (удлинение полинуклеотидной цепи) и терминацию (окончание процесса). Синтез мРНК начинается с обнаружения ферментом РНК-полимеразой особого участка нуклеотидной последовательности промотора.
После присоединения, РНК полимераза расщепляет двойную цепочку ДНК и на одной из ее цепей по принципу комплементарности осуществляется синтез иРНК (рис. 8). В связи с тем, что РНК-полимераза способна собирать полинуклеотид лишь от 5´к 3´ концу матрицей для транскрипции может служить только одна из цепей ДНК, которая обращена к ферменту своим 3´концом (3´ - 5´). Такая цепь называется кодогенной. Построение комплементарной иРНК будет идти до тех пор, пока не встретится специальная последовательность нуклеотидов - терминатор. На этом участке синтезированная мРНК отделяется от материнской ДНК. Фрагмент молекулы ДНК включающий промотор, транскрибируюмую часть, терминатор образуют единицу транскрипции – транскриптон.

Рис. 8. Схема синтеза мРНК
Трансляция заключается в синтезе полипептида на рибосоме, при котором в качестве матрицы используется молекула мРНК. Как и в случае транскрипции, процесс трансляции можно условно подразделить на три основные стадии - инициацию, элонгацию и терминацию (рис. 9). В процессе трансляции участвуют также молекулы тРНК, функция которых состоит в транспортировке аминокислот из цитоплазмы к рибосомам. Транспортные РНК приносят аминокислоты в большую субъединицу рибосомы.
В синтезе первичной структуры белка принимают участие рибосомы. В рибосомах выделяют два участка аминоацильный (А) - центр узнавания аминокислот. В этом участке располагается аминоацил тРНК, несущая определенную аминокислоту и пептидальный (П) центр образования пептидной связи. Здесь располагается обычно тРНК, нагруженная цепочкой аминокислот. Образование А и П участка обеспечивается обеими субчастями рибосом.
|
|
|
Фаза инициации заключается в объединении двух субъединиц рибосом на определенном участке мРНК и присоединении к ней первой аминоацил тРНК. Этим задается рамка считывания информации. Функциональная особенность П-участка состоит в том, что он может быть занят только инициирующей аминоацил-тРНКс антикодоном УАГ, который у эукариот несет аминокислоту метионин, а у прокариот - формилметионин. К концу фазы инициации П-участок занят аминоацил тРНК, в участке А располагается следующий за стартовым кодон.

Рис. 9. Начальные этапы трансляции: А – инициирующий комплекс, Б – элонгация.
Фаза элонгации – тРНК с аминокислотой подходит к аминоацильному центру рибосомы, если антикодон т-РНК комплементарен кодону и РНК, то происходит временное присоединение тРНК саминокислотой к кодону иРНК
После этого рибосома передвигается на один кодон-иРНК и т-РНК с аминокислотой перемещается в пептидальный центр, а к освободившемуся аминоацильному центру приходит новая т-РНК. С помощью ферментов находящихся в рибосоме устанавливается пептидная связь между аминокислотами. Одновременно разрушается связь между первой аминокислотой и ее т-РНК, а также т-РНК и и-РНК. Т-РНК уходит из рибосомы. Рибосома опять перемещается на один триплет и процесс повторяется. Считывание информации идет в одном направлении 5´- 3´. В синтезируемой молекуле полипептида аминокислоты располагаются в строгом соответствии с порядком кодирующих триплетов (коллинеарность). Сборка пептидной цепи осуществляется с достаточно большой скоростью, зависящей от температуры. У порокариот при температуре 37o к полипептиду добавляется в одну секунду до 12-17 аминокислот, у эукариот 2.
Фаза терминации. Терминация трансляции связана с вхождением одного из трех известных стоп кадонов м РНК (УАА, УАГ, УГА) в А-участок рибосомы. Так как такой триплет не несет информации о какой либо аминокислоте, но узнается соответствующими белками терминации, то процесс синтеза полипептида прекращается и он отсоединяется от матрицы мРНК. После выхода из рибосомы свободный 5´конец мРНК может вступить в контакт со следующей рибосомой полисомной группы, инициируя синтез идентичного полипептида.
Таким образом, перенос генетической информации осуществляется по схеме ДНК – РНК – белок – признак. Это сложившееся представление о переносе генетической информации принято называть центральной догмой молекулярной биологии (рис. 10).

Рис.10. Центральная догма молекулярной биологии.
Наряду с этим наиболее распространенным направлением переноса известна и другая форма реализации генетической информации – специализированный перенос. Эта форма переноса обнаружена у РНК-содержащих вирусов. В этом случае наблюдается процесс, получивший название обратной транскрипции. При обратной транскрипции вирусная РНК проникает в клетку хозяина и служит матрицей для синтеза комплементарной ДНК. ДНК синтезируется с помощью фермента обратной транскриптазы (ревертазы), кодируемой вирусным геномом. В дальнейшем возможна реализация информации синтезированной вирусной ДНК в обычном направлении. Следовательно, специализированный перенос генетической информации осуществляется по схеме РНК – ДНК – РНК - белок.
Белки
Белки высокомолекулярные полимерные соединения мономерами, которых являются аминокислоты. В состав молекулы каждого белка входят определенные аминокислоты в свойственном этому белку количественном соотношении и порядке расположения в пептидной цепи.
Структуры белка: первичная, вторичная, третичная, четвертичная. Белки выполняют следующие функции:
· ферментативную;
· структурную;
· рецепторную;
· транспортную;
· защитную;
· двигательную;
· регуляторную;
· энергетическую.
Таблица 2
Классификация генов
| Участок ДНК | Функция |
| Структурные гены | Имеются у про- и эукариот и содержат информацию о строении белков, выполняющих, специфические общеклеточные функции, а также информацию о тРНК и рРНК |
| Экзоны | Информативные участки генов |
| Оператор | Расположен у прокариот перед структурными генами и служит для связи с белком-репрессором |
| Промотор | Служит для связи с ферментом РНК-полимеразой. У прокариот представлен ТАТАААТ последовательностью (блок Пробнова), а у эукариот ТАТА-блоком или ЦАТТ – блоком, которые расположены на разных расстояниях от стартовой точки транскрипции |
| Терминатор | Служит для прекращения роста цепи РНК и его освобождения от матрицы ДНК. У большинства прокариот в связи с наличием факторов антитерминации транскрипция продолжается за пределами терминатора, что ведет к образованию полицистронной мРНК. У эукариот терминатор останавливает движение РНК-полимеразы и в итоге образуется моноцистронная мРНК |
| Сенсорный ген | Служит у эукариот для восприятия химического индукционного сигнала |
| Ген регулятор | У прокариот содержит информацию о структуре белков-репрессоров, регулирующих работу других генов |
| Гены модуляторы: | Обнаружены только у эукариот |
| Энхансеры | Усиливают транскрипцию |
| Сайленсеры Мутаторы Антимутаторы | Тормозят транскрипцию Способствуют мутациям других генов Снижают мутагенный эффект |
| Неинформативные участки ДНК: Интроны Спейсеры Сателлитная ДНК | Имеются в составе генов эукариот и не содержат информацию о белке, кодируемым данным геном Некодирующие последовательности, разделяющие структурные гены Участвует в коньюгации хромосом эукариот, может содержать мобильные элементы. |
Таблица 3
Генетическая характеристика прокариот и эукариот
| Признаки | Прокариоты | Эукариоты |
| Количество генов | 4 тыс. (E. coli) | Около 30 тыс. (человек) |
| Количество ДНК | 4 млн. пар нуклеотидов | 3-7 млрд. пар нуклеотидов |
| Кодирующие последовательности ДНК | Более 90% | Менее 10 % |
| Связь ДНК с гистонами | Отсутствует | Формирует нуклеосомы |
| Укладка ДНК | Кольцевая содержит 100 петель по 40 тыс. пар нуклеотидов | Линейная с замкнутыми в теломеры концами, имеет 4 уровня спирализации |
| Количество репликонов | Один | 50 тыс. |
| Активно работающие участки | Более 90 % генов | Менее 10 % генов |
| Процессинг | Отсутствует | Осуществляется при переходе пре- м РНК из ядра в цитоплазму |
| Регуляция транскрипции | Опероннная | Сложная, каскадная |
Самостоятельная работа.
|
|
|
