 |


Физиология центральной нервной системы
|
|
|
|
Физиология центральной нервной системы (ЦНС) — важнейший стержневой раздел физиологической науки, так как ЦНС влияет на все процессы в организме и в то же время сама подвергается воздействию каждого из них.
ЦНС объединяет все процессы, происходящие в организме, определяет поведение животного и его взаимоотношения с окружающей средой. Развиваясь в процессе этих взаимодействий более, чем какая-либо другая система организма, она играет важнейшую роль в эволюционном развитии всех его функций.
Благодаря усовершенствованию старых и применению новых методов исследования физиология ЦНС обогатилась многими новыми фактами, которые открыли путь для перехода от гипотез относительно механизмов деятельности мозга к прямому и точному их изучению. Благодаря использованию в нейрофизиологии таких новых экспериментальных методов, как микроэлектронная регистрация потенциалов, электронная микроскопия, микробиохимические исследования в сочетании с математическим и физическим моделированием, их компьютерной обработке, получены обстоятельные данные о распространении нервного импульса, трансмембранных ионных токах, природе синаптической передачи, роли нейроглии в регуляции состава внутримозговой межклеточной среды и в генерации длительных колебаний электрических потенциалов мозга и др.
ЦНС присущи такие функции, как восприятие афферентных (центростремительных) импульсов, возникающих при раздражении рецепторов, расположенных во всех органах и тканях; анализ и синтез этих раздражений и формирование потоков эфферентных (центробежных) импульсов, либо вызывающих, либо прекращающих деятельность периферических органов, либо поддерживающих их тонус. ЦНС обеспечивает индивидуальное приспособление организма к окружающей среде соответственно его потребностям, наиболее совершенное координирование деятельности всех систем, органов и тканей.
|
|
|
Эти сложнейшие и жизненно важные процессы осуществляются с помощью нервных клеток — нейронов, специализированных на восприятии, хранении и передаче информации и объединенных в специфически организованные нейронные цепи и центры, составляющие различные функциональные системы мозга.
НЕЙРОНЫ И СИНАПСЫ
В основе современного представления о структуре и функции ЦНС лежит нейронная теория. Нервная система построена из двух типов клеток: нервных и глиальных, причем число последних в 8...9 раз превышает число нервных. Однако именно нейроны обеспечивают все многообразие процессов, связанных с передачей и обработкой информации.
Таким образом, структурно-функциональная единица ЦНС — нервная клетка, или нейрон. Отдельные нейроны, в отличие от других клеток организма, действующих изолированно, «работают» как единое целое. Их функции заключаются в передаче информации (в форме сигналов) от одного участка нервной системы к другому, в обмене информацией между нервной системой и различными участками тела. При этом передающие и принимающие нейроны объединены в нервные сети и цепи.
В нервных клетках происходят сложнейшие процессы обработки информации. С их помощью на внешние и внутренние раздражения формируются ответные реакции организма — рефлексы.
Нейроны обладают рядом признаков, общих для всех клеток тела, независимо от местонахождения и функций. Любой нейрон имеет плазматическую мембрану, определяющую границы индивидуальной клетки. Нейрон взаимодействует с другими нейронами или улавливает изменения в локальной среде с помощью мембраны и заключенных в ней молекулярных механизмов. Стоит отметить, что мембрана нейрона значительно прочнее, чем других клеток.
|
|
|
Все, что заключено внутри плазматической мембраны (кроме ядра), называется цитоплазмой. Здесь содержатся цитоплазмати-ческие органеллы, необходимые для существования нейрона и выполнения им своей работы. Митохондрии обеспечивают клетку энергией, используя сахар и кислород для синтеза специальных высокоэнергетических молекул, расходуемых клеткой по мере надобности. Микротрубочки — тонкие опорные структуры — обеспечивают поддержание определенной формы нейрона. Сеть внутренних мембранных канальцев, с помощью которых клетка распределяет химические вещества, необходимые для ее функционирования, называется эндоплазматическим ретикулумом.
Различают два вида эндоплазматического ретикулума: шероховатый и гладкий. Мембраны шероховатого (гранулярного) усеяны рибосомами, необходимыми клеткам для синтеза секретируемых ими белковых веществ. Обилие элементов шероховатого ретикулума в нейронах характеризует их как клетки с весьма интенсивной деятельностью. Другой вид плазматического ретикулума — гладкий, называемый также аппаратом Гольджи, упаковывает вещества, синтезированные клеткой, в специальные «мешочки», построенные из мембран гладкого ретикулума. Задача этой органеллы нейрона заключается в переносе секретов к поверхности клетки.
35 — 3389

|
Рис. 13.1. Схематическое изображение нейрона (пропорции изменены):
/ — девдриты; 2— тело клетки; 3— аксонный холмик; 4— аксон; 5 — коллатераль аксона; 6 — миелиновая оболочка; 7— шваннов-ская оболочка; 8— ядро шванновской клетки; 9— перехват Ран-вье; 10— концевые разветвления аксона, лишенные миелиновой оболочки
В центре цитоплазмы находится ядро, в котором, как и у всех клеток с ядрами, содержится генетическая информация, закодированная в химической структуре генов. В соответствии с этой информацией полностью сформированная клетка синтезирует специфические вещества, которые определяют ее форму, химизм и функцию. Однако в отличие от большинства других клеток тела зрелые нейроны не могут делиться. Поэтому генетически обусловленные химические элементы любого нейрона должны обеспечивать сохранение и изменение функций на протяжении всей его жизни. В крупных нейронах ядро занимает четверть или треть их тела. Входящие в его состав ядрышки участвуют в снабжении клетки рибонуклеиновыми кислотами и белками (в мотонейронах, например, при двигательной активности животного ядрышки значительно увеличиваются в размерах).
|
|
|
Вместе с тем нейроны в отличие от других клеток организма кроме тела (сомы) имеют отростки (рис. 13.1). Многочисленные чувствительные короткие древовидно-разветвленные отростки — дендриты (гр. дерево) служат своеобразными входами, через которые сигналы поступают в нервную клетку. Они имеют шероховатую поверхность, обусловленную небольшими утолщениями — шипиками, словно бусинки, нанизанными на дендрит. Благодаря этому увеличивается поверхность нейрона и максимально повышается сбор информации.
Выходом нейрона является отходящий от гена длинный гладкий отросток — аксон (от гр. axis — ось), который передает нервные импульсы другой нервной клетке или рабочему органу. Аксоны многих нейронов покрыты миелиновой оболочкой, образованной шванновскими клетками, многократно (10...65 слоев) «намотанными» подобно изоляционной ленте вокруг ствола аксона. Однако муфты шванновских клеток, надетые на аксон, не соприкасаются друг с другом. Между ними остаются узкие щели — перехваты Ранвье, благодаря которым нервное волокно непосредственно соприкасается с внеклеточной жидкостью. Поэтому в нервной системе млекопитающих животных волна нервного им-
пульса распространяется не плавно, а скачками (сальтаторно) от одного перехвата к другому, что весьма ускоряет процесс продвижения импульса.
Начальная часть аксона — начальный сегмент в месте выхода из тела клетки (область «аксонного холмика») лишена миелиновой оболочки, а его мембрана обладает высокой возбудимостью. Поэтому ее называют пусковой зоной, так как именно отсюда начинается возбуждение нейрона.
Для осуществления внутримозговых связей нужны очень длинные отростки; аксоны выходят за пределы ЦНС и следуют к мышцам, железам, внутренним органам. Собранные в пучки, они образуют нервы. Длина аксонов, идущих на периферию, зависит от размера животного: у крупной собаки — 0,5 м, у лошади — более 1, у жирафа и слона —3, а у гигантского кальмара — 8 м.
|
|
|
Если нейрон образует выходные связи с большим числом других клеток, то его аксон может многократно ветвиться, чтобы сигналы могли дойти до каждой из них: количество таких разветвлений (термиполей) огромно — от 1000 до 10000 и более. Кроме того, аксон способен отдавать дополнительные ветви — коллатерали, по которым возбуждение уходит далеко в сторону от магистрального пути. Небезынтересен следующий факт — отростки, разобщенные с телом клетки, долго существовать не могут и погибают. Тело клетки, напротив, регенерирует их. Конечно, это относится только к центральной части отростка. Иногда процессы регенерации отростков идут с огромной скоростью: до 30 мк в 1 мин.
Следует отметить, что именно из-за наличия отростков нейроны как клетки были открыты позже других клеток, так как в поле зрения микроскопа нейрон со всеми своими отростками поместиться не мог. Поэтому первоначально самим клеткам не придавали должного значения, рассматривая их как утолщение среди множества отростков.
Форма нейрона, размер, расположение отростков разнообразны и зависят от функционального назначения. Каждый отдельный нейрон уникален и неравноценен себе подобным в отличие от других клеток организма. Размеры нейронов весьма вариабельны: самые крупные в десятки и сотни раз больше самых мелких. К примеру, поперечник зернистых клеток мозжечка составляет 7 мк, а моторных нейронов спинного мозга — 70 мк.
Плотность расположений нейронов в некоторых отделах ЦНС очень велика. Так, в коре больших полушарий она составляет 40 000 клеток в 1 мм3. Считается, что мозг человека и высокоорганизованных животных содержит приблизительно десятки миллиардов нейронов.
Классификация нейронов. Современная нейробиология предлагает две классификации нейронов.
35*
Согласно первой все нервные клетки делятся на три типа: афферентные (центростремительные, сенсорные, чувствительные), эфферентные (центробежные) и промежуточные (вставочные, контактные, ассоциативные, интернейроны).
Афферентные нейроны передают импульсы (информацию) от рецепторов в ЦНС. Тела их расположены вне ЦНС — в спинномозговых или черепно-мозговых ганглиях (рядом с головным и спинным мозгом). Афферентный нейрон имеет псевдоуниполярную форму, т. е. оба его отростка выходят из одного полюса клетки. Один из отростков направляется на периферию, где заканчивается рецептором (аксоноподобный дендрит), а другой — в ЦНС (истинный аксон). К афферентным нейронам также относятся нервные клетки, аксоны которых составляют восходящие пути головного и спинного мозга.
|
|
|
Эфферентные нейроны работают в центробежном режиме, т. е. они связаны с передачей нисходящих импульсов от вышерасположенных этажей нервной системы к нижерасположенным. Например, от коры к спинному мозгу или от спинного мозга к рабочим органам. Для эфферентных нейронов характерны разветвленная сеть дендритов и один длинный аксон. Необходимо отметить, что число эфферентных нейронов в 4...5 раз меньше афферентных.
Промежуточные нейроны, как правило, более мелкие клетки, осуществляющие связь между различными нейронами (в частности, афферентными и эфферентными). Они передают нервные импульсы в различных направлениях (горизонтальном, вертикальном) по ЦНС. Благодаря многочисленным разветвлениям аксона промежуточные нейроны могут одновременно возбуждать большое число других нейронов. В ЦНС преобладают промежуточные нейроны (рис. 13.2).
По второй классификации все нейроны делятся на возбуждающие и тормозные.
Нейроглия. Бесспорно, что нервные клетки — главные труженики мозга, но их в мозге значительно меньше, чем других клеток. Оказывается, для осуществления процессов нормального питания и обмена веществ нейронов необходимы многочисленные клетки нейроглии (глиальные клетки). В мозговой ткани они заполняют все пространство между нейронами: в коре больших полушарий их примерно в 5 раз больше, чем нейронов.
Различают три типа клеток нейроглии: астроциты (звездообразные, с большим количеством отростков); олигодендро-циты (округлые или многоугольные с малым количеством отростков); микроглия (мелкие клетки разнообразной формы).
Клетки нейроглии плотно окружают всю капиллярную сеть в мозговой ткани; свободна от них лишь незначительная часть поверхности сосудов (около 15 %). Выросты глиальных клеток могут

Рис. 13.2. Типы нервных клеток:
/— нейроны; 2— промежуточные нейроны; 3— эфферентные нейроны
располагаться с одной стороны на нейроне, с другой — на кровеносных сосудах, что указывает на их участие в передаче питательных веществ и кислорода из крови в нервную клетку. Экспериментально доказано, что нейроглия активно поддерживает функционирование нейрона. Например, при длительном возбуждении высокое содержание белка и нуклеиновых кислот в нейроне поддерживается за счет клеток глии, в которых уровень этих веществ соответственно падает. В процессе же восстановления после работы запасы белка и нуклеиновых кислот сначала нарастают в клетках этой глии, а лишь затем в цитоплазме нейрона. Интересно отметить, что нейроглиальные клетки весьма мобильны и могут перемещаться в направлении к более активным нейронам. Таким образом, в случае необходимости компенсируется доставка питательных веществ и кислорода к активно «работающим» нейронам.
Клетки нейроглии, предположительно, выполняют функцию своеобразной гидродинамической подушки, предохраняющей чувствительные и нежные образования нейронов от различных физи-
ческих воздействий. В последнее время появились сообщения об участии этих клеток в условно-рефлекторной деятельности мозга и механизмах памяти.
Таким образом, система нейрон — нейроглия постоянно находится в состоянии гибкого ритмически колеблющегося равновесия. Но в этих процессах, происходящих в мозгу, безусловно, преобладают нейроны, используя нейроглию для поддержания своего функционирования.
Синапсы в ЦНС. Деятельность мозга невозможна без взаи
модействия нейронов друг с другом. Трудность в передаче воз
буждения заключается в том, что каждая нервная клетка, одетая
в собственную оболочку, окруженная нейроглией и закутанная
слоями миелина, представляет собой маленькое самостоятельное
образование. *
Для обмена информацией необходимы специальные устройства. Эти устройства — места контакта двух нейронов — назвал синапсами (от гр. synapsis — связь, соединение, смыкание) в конце XIX в. английский физиолог Чарльз Шеррингтон. Правда, в то время еще не было известно, что нервные клетки обмениваются между собой с помощью химических передатчиков. Такие сигналы в дальнейшем были названы химическими (они преобладают в мозге млекопитающих). Кроме них в ЦНС, в основном у низших животных, встречаются электрические (эфапсы) и смешанные синапсы. Химические синапсы состоят из пресинапти-ческой и постсинаптической мембран (рис. 13.3). Пресинапти-ческая мембрана содержит небольшие поры, способные пропускать медиатор, который как бы упакован в крохотных синапти-ческих пузырьках диаметром 20...60нм, локализованных вблизи мембраны. Выделение медиатора становится возможным благодаря тому, что синаптические пузырьки «сдвигаются» нервным импульсом в область так называемой активной или оперативной зоны. Чем больше нервных импульсов проходит через синапс, тем больше пузырьков скапливается в этой зоне и прикрепляется к пресинаптической мембране. В результате этого облегчается выделение медиатора при последующих нервных импульсах.
В каждом пузырьке пресинаптической мембраны содержится несколько тысяч молекул медиатора. При этом некоторые поры мембраны прикрыты неплотно и пузырьки постоянно (по одному в секунду) изливаются в синаптическую щель. Однако высвобождение такой ничтожной порции медиатора остается без ответа, так как не хватает критической массы вещества для осуществления тех процессов, которые должны развиваться.
Известны два типа химических синапсов, различающихся по характеру процессов, развивающихся на постсинаптической мембране: возбуждающие и тормозные.
Рассмотрим механизм работы возбуждающего синап-с а. Во время действия нервного импульса на нервное окончание из пузырьков выделяется порция (квант) медиатора и поступает через поры пресинаптической мембраны в синаптическую щель. Предполагается, что только благодаря свободной диффузии молекулы медиатора могут пересечь это пространство за считанные 1...2 с-3 (миллисекунда). Как только медиатор вступит в контакт с белками-рецепторами в области постсинаптической мембраны, последняя становится проницаемой для ионов Na+. Эти ионы лавинообразно устремляются в клетку и переносят положительный заряд на внутреннюю поверхность мембраны. После деполяризации постсинаптической мембраны возникает возбуждающий постсинаптический потенциал (ВПСП). При этом регистрируется небольшое колебание мембранного потенциала с амплитудой до 10 мВ. В дальнейшем он перерастает в потенциал действия. Необходимость в медиаторе отпадает, и он разрушается специфическим ферментом синапса.
Существуют различия в строении и функционировании синапсов (рис. 13.4):
синаптическая щель тормозного синапса уже и составляет 20 нм (у возбуждающего — 30 нм);
постсинаптическая мембрана тормозного синапса более толстая и плотная;
пресинаптическая мембрана тормозного синапса содержит меньше пузырьков медиатора;
медиаторами в тормозных синапсах являются глицин, гамма-аминомасляная кислота (ГАМК), иногда — ацетилхолин.
Для включения в работу тормозного синапса необходим также импульс возбуждения. При достижении импульсом пресинаптической мембраны в синаптическую щель выбрасывается порция тормозного медиатора. Действуя как химический раздражитель на мембранные рецепторы, тормозной медиатор незначительно изменяет проницаемость мембраны в основном для ионов К+ и С1_. Поскольку ионов К+ больше внутри клетки, а ионов С1~ — в межклеточном пространстве, они двигаются в противоположные направления: К* — наружу, С1- — внутрь, что увеличивает поляризованность мембраны (мембранный потенциал).


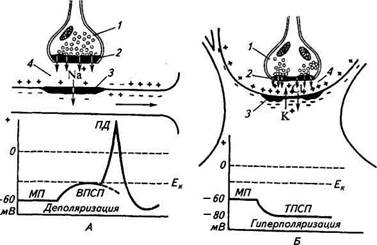
Рис. 13.4. Схема функции возбуждающего (А) и тормозящего (£) синапсов:
1 — пузырьки медиатора; 2— пресинаптическая мембрана; 3 — постсинаптическая
мембрана; 4— синаптическая щель; ^ — критический уровень деполяризации-
МП— местный потенциал; ВПСП— возбуждающий и ТПСП — тормозной пост-
синаптические потенциалы; ПД— потенциал действия
При этом регистрируется положительное колебание с амплитудой около 5 мВ — тормозной постсинаптический потенциал (ТПСП). Продолжительность ТПСП (как и ВПСП) всего несколько миллисекунд, но в течение этого времени никакое возбуждение на данном конкретном участке постсинаптической мембраны невозможно. В этом и заключается механизм действия тормозного синапса. Необходимо отметить, что в некоторых нейронах головного мозга продолжительность постсинаптического потенциала довольно значительная: ВПСП — до 80 с-3, ТПСП — до 100 с~3.
Принимая во внимание то, что возбуждение любого нейрона обязательно сопровождается возникновением биоэлектрических потенциалов, возникает вопрос: почему же эволюция пошла по более сложному и медленному пути проведения возбуждения — через химический синапс? Казалось бы, все очень просто: ведь электричество — отличный раздражитель, и для передачи возбуждения с одной нервной клетки на другую достаточно наличия электрического синапса. Однако мозг не способен различать нужное напряжение электрического тока, ведь нейрон работает по закону «все или ничего», т. е. или «молчит», или дает стандартный ответ. Там, где возбуждение обязательно передается от клетки к клетке, этот принцип можно было бы применить. Но в мозгу чаще всего возникает такая ситуация, когда нервная клетка вовсе не отвечает немедленным возбуждением. В этом и состоит работа ней-
ронов: постоянно анализировать, достаточно ли основательна поступающая информация. Именно поэтому нервные клетки высших животных обмениваются между собой информацией посредством химических веществ, которые легко дозируются.
В мозговой деятельности участвуют несколько медиаторов: нор-адреналин, донамин, сербонин вызывают возбуждение; глицин и гамма-аминомасляная кислота (ГАМ К) — торможение; ацетил-холин, универсальный медиатор, — возбуждение и торможение. Каждому синапсу соответствует только один медиатор.
По другой классификации различают следующие синапсы:
1. Аксоносоматические — между аксоном одного и телом другого нейрона.
2. Аксодендритические — между аксоном одного и дендритом другого нейрона.
3. Аксо-аксональные — между аксонами двух нейронов.
Возможны и другие варианты. При этом возбуждающими, как
правило, бывают аксодендритические, а тормозными — аксоносоматические и аксо-аксональные синапсы.
В ЦНС высокоорганизованных животных доля электрических синапсов (эфапсов) незначительна. Передача возбуждения через эти синапсы называется эфаптпической. Пресинаптическая мембрана в таких синапсах сообщается с постсинаптической посредством миниатюрных мостиков, т. е. мембраны не разделены щелью. Потенциал действия, достигнув пресинаптической мембраны, преобразуется в градуальный ток, «стекающий» на пост-синаптическую мембрану, где возрождается как новая волна возбуждения. Таким путем возбуждение проводится через синапс почти без задержки. Особенность эфапсов заключается в их способности передавать возбуждение в двух противоположных направлениях, что невозможно в химических синапсах (рис. 13.5).

|
| Рис. 13.5. Схема синапсов с химическими (А), электрическими (Б) и смешанными (В) механизмами передачи: |
| / — пресинаптическая и 2—постсинаптическая мембраны; 3 — синаптическая щель |
В мозговой ткани встречаются и смешанные синап-с ы. Такой синапс частично может проводить возбуждение как эфапс или с помощью медиатора.
Функции нейронов. Нейрон обладает следующими важнейшими функциями: рецепторной — восприятие различных раздражений; интегративной — переработка информации; эффек-торной — передача нервных импульсов на другие нейроны или органы-эффекторы.
Рецепторная (воспринимающая) функция нейрона заключается в передаче возникающих в рецепторах возбуждений в нервную систему через определенные участки нейрона, находящиеся в области синаптических контактов. Ответ нейронов на раздражение проявляется изменением величины мембранного потенциала.
Чем больше синапсов на нервной клетке, тем больше различных раздражений она воспринимает и, следовательно, тем шире спектр влияний на деятельность клетки и возможность ее участия в разнообразных реакциях организма. Тело нейрона и значительная часть дендритов (50 %) буквально усыпаны синапсами. Особенно густо синапсы покрывают среднюю часть и окончание дендритов, причем многие контакты расположены на шипиках. Так, в мотонейронах спинного мозга и пирамидных клетках коры поверхность дендритов в 10...20 раз больше поверхности тела клетки.
Рецепторную функцию выполняют афферентные нейроны, имеющие большое число различных синапсов (возбуждающих и тормозных), способных к сложным процессам переработки поступающей в ЦНС информации.
Интегративная функция нейрона — это общее изменение его мембранного потенциала в результате сложного взаимодействия (интеграции) местных ВПСП и ТПСП всех многочисленных активированных синапсов на его теле и дендритах. На мембране нейрона происходит процесс суммирования положительных и отрицательных колебаний потенциала.
В конечном итоге реакция нервной клетки определяется суммой всех синаптических импульсов. Так, преобладание тормозных синаптических воздействий приводит к гиперполяризации мембраны и торможению деятельности нейрона. При сдвиге же мембранного потенциала в сторону деполяризации возбудимость нервной клетки повышается. Однако электрический разряд нейрона возникает лишь тогда, когда изменения мембранного потенциала достигают порогового значения — критического уровня деполяризации (около 10 мВ).
Как уже отмечалось ранее, в нейронах (прежде всего крупных) возбудимость различных участков мембраны неодинакова. В области начального сегмента нервной клетки (аксонного холмика и следующей за ним немиелизированной части аксона) находится низкопороговая зона, в которой мембрана нейрона обладает более высокой возбудимостью, чем на других участках клетки (так, порог возбудимости данной области составляет 10 мВ, а области дендрита и других районов сомы — 20...30 мВ). Именно в этой об-
ласти с момента достижения критического уровня деполяризации начинается лавинообразное движение ионов Na+ в клетку и регистрируется потенциал действия (ПД). Поэтому эту зону принято называть пусковой зоной нейрона.
В ПД нервной клетки, так же как и в клетках других возбудимых тканей (мышечной, железистой), различают кратковременную высоковольтную часть, или пик, и длительные низкоамплитудные колебания — это следовые потенциалы. При этом необходимо отметить, что амплитуда пика ПД большинства мотонейронов колеблется от 80 до 100 мВ, а его длительность составляет 1,5 с-3.
Эфферентная функция нейрона заключается в проведении возбуждения от тела нейрона по аксону к другой нервной клетке или рабочему органу. В формировании окончательного эфферентного сигнала решающую роль играют синапсы, расположенные ближе к возбудимой низкопороговой зоне на теле клетки (аксоносоматические синапсы). Импульсы, проходящие через эти синапсы, как правило, вызывают ответный разряд нейрона, а импульсы, действующие через отдаленные (аксодендритические) синапсы, могут привести лишь к подпороговым изменениям его возбудимости. Таким образом, аксоносоматические синапсы создают в основном условия для ответа нервной клетки на раздражение, но будет ответ или нет обусловливается характером воздействий, поступающих через аксодендритические синапсы от других нервных центров.
Таким образом, складываются адекватные реакции, зависящие от многих раздражений, действующих на организм в конкретный момент времени. В результате всего этого осуществляется тонкое приспособление организма к меняющимся условиям внешней и внутренней среды.
Физиологические показатели функционального состояния нейрона. Функциональное состояние нейрона непостоянно и может существенно изменяться. Физиологическими показателями этого состояния являются возбудимость и лабильность.
Возбудимостью нейрона называют его способность отвечать на синаптическое воздействие потенциалом действия. Возбудимость определяется соотношением "двух параметров — мембранного потенциала и критического уровня деполяризации. Надо отметить, что в нормальных условиях деятельности нейрона критический уровень деполяризации относительно стабилен (=10 мВ), поэтому возбудимость нервной клетки в основном зависит от величины мембранного потенциала. Обычно потенциал покоя мембраны нейрона в норме составляет около 70 мВ, но он может быть уменьшен (явление деполяризации) или увеличен (явление гиперполяризации).
Как уже неоднократно отмечалось, для появления ПД при нормальном мембранном потенциале необходим вначале оптимальный сдвиг заряда мембраны в сторону деполяризации на 10 мВ
(амплитуда порогового ВПСП) — эта величина называется порогом возбудимости нейрона.
Посмотрим, как меняется возбудимость нейрона при снижении его мембранного потенциала (это наблюдается в активно работающей нервной клетке или при слабых синаптических воздействиях, когда появляются подпороговые ВПСП). При этом разница между уровнем мембранного потенциала и критическим уровнем деполяризации уменьшается. Значит, снижается и амплитуда порогового ВПСП. В итоге для появления ответного ПД требуется меньшая сила раздражителя. Возбудимость нейрона в этом случае повышена.
При понижении возбудимости нервной клетки разница между уровнем мембранного потенциала и критическим уровнем деполяризации возрастает, и для возникновения ПД требуется значительно более сильное раздражение (большая амплитуда порогового ВПСП).
Возбудимость нейрона при его возбуждении изменяется с той же закономерностью, что и в других клетках возбудимых тканей. В момент появления высоковольтной части — пика (спайка) нейрон не может ответить на раздражение новым ПД, так как он абсолютно невозбудимый (абсолютная рефрактерность). Эта фаза длится примерно 0,5 с-3. Затем, в период реполяризации мембраны, возбудимость нервной клетки постепенно восстанавливается до исходного уровня (относительная рефрактерность). Следующей фазой является экзальтация (повышение уровня возбудимости в сравнении с исходным состоянием), и, наконец, во время следовой гиперполяризации наступает фаза субнормальности (незначительное понижение уровня возбудимости).
Знание закономерностей изменения возбудимости нейрона при возбуждении имеет большое значение для понимания особенностей его деятельности в различных ситуациях. Так, большинство нейронов промежуточного мозга и коры больших полушарий после очередного разряда импульсов (из-за соответствующей продолжительности фаз абсолютной и относительной рефрактерное™) неактивно в течение примерно 100 с-3. Следовательно, наиболее удобный ритм их спонтанной активности — около 10 разрядов в 1 с, чем определяется частота электрических колебаний на поверхности мозга в состоянии покоя.
Лабильность — это скорость протекания элементарных реакций, лежащих в основе возбуждения нервной клетки. Разные по функциям и размерам нейроны обладают различными величинами лабильности: даже в пределах одной нервной клетки лабильность ее разных структур (сомы, дендритов, начального сегмента аксона) резко различается. Величина лабильности нейрона, так же как и возбудимости, определяется уровнем его мембранного потенциала. Лишь при определенной величине мембранного потенциала достигается оптимальный уровень возбудимости и лабиль-
ности нервной клетки, а также наиболее высокий уровень ее ритмической активности. Это обстоятельство, бесспорно, является важным условием для передачи информации в ЦНС и осуществления целесообразных реакций. Например, максимальный ритм мотонейронов спинного мозга (до 100 импульсов в секунду) отмечается при средней, оптимальной величине мембранного потенциала — 55 мВ, т. е. при повышенной возбудимости мотонейронов.
В естественных условиях жизнедеятельности частота разрядов мотонейрона обычно не превышает 50...60 имп/с, но иногда она может повышаться до 100...300 имп/с. Это бывает при мощном влиянии вышерасположенных отделов ЦНС и естественном снижении мембранного потенциала. Однако разряды с такой частотой очень кратковременны. Они сменяются более медленным и устойчивым ритмом активности.
В мелких же промежуточных нейронах спинного мозга частота разрядов в начальные моменты активации может достигать 600...800 и даже 1000 имп/с. Это необходимо для резкого и быстрого воздействия этих клеток на мотонейроны.
Многочисленные раздражения внешней и внутренней среды, вызывающие афферентную импульсацию, повышают возбудимость и лабильность нейронов, их способность к стабильной ритмической активности, а бездеятельное же состояние понижает их, ухудшая функциональное состояние нервной клетки.
Активная деятельность нейрона вызывает не только функциональные изменения. Она ведет к значительным структурным и биохимическим перестройкам его различных элементов (например, к изменениям в синаптическом аппарате нервных клеток). Так, эксперименты с тренировками животных (нагрузки на различные группы мышц) показали, что средняя величина (диаметр) синапсов на мотонейронах спинного мозга достоверно возрастает, примерно на 35 %. С увеличением размеров синапсов улучшается проведение нервных импульсов в ЦНС.
Активное участие корковых нейронов в условно-рефлекторной деятельности способствует развитию шипикового аппарата на дендритах и усиливает межнейронные взаимосвязи в коре больших полушарий. Подавление деятельности коры различными фармакологическими препаратами, наоборот, приводит к ухудшению рецепторной функции нейронов — нарушению синаптических контактов (исчезновению шипиков) и даже деформации дендритов.
При усилении афферентной импульсации, поступающей к нейрону, в синапсах происходит усиление синтеза медиатора и соответственно увеличение числа синаптических пузырьков, а также их интенсивное перемещение в оперативную зону пресинапти-ческой мембраны. Необходимо отметить, что на фоне усиленной двигательной активности могут увеличиваться даже размеры эф-фекторных нейронов.
 Таким образом, все структурные, биохимические и функциональные изменения способствуют осуществлению нервными клетками их сложных функций. Это лежит в основе совершенствования различных поведенческих реакций целостного организма, в том числе двигательной деятельности и психических процессов.
Таким образом, все структурные, биохимические и функциональные изменения способствуют осуществлению нервными клетками их сложных функций. Это лежит в основе совершенствования различных поведенческих реакций целостного организма, в том числе двигательной деятельности и психических процессов.
РЕФЛЕКТОРНАЯ ДЕЯТЕЛЬНОСТЬ
Рефлекс — это ответная реакция организма на раздражение при участии нервной системы.
В основе рефлекторного процесса лежит рефлекторная дуга — комплекс специфически организованных нервных элементов, взаимодействие которых необходимо для осуществления рефлекторного акта. Дуга простого рефлекса состоит из рецепторов, чувствительного, или афферентного, нейрона, промежуточного нейрона, эфферентного нейрона и эффектора (рис. 13.6).
Афферентная часть представлена теми нервными элементами, которые формируют и проводят в центральном направлении нервные импульсы, необходимые для деятельности всей рефлекторной дуги. Возникновение афферентных импульсов связано с активацией специфических рецепторов; совокупность рецепторов, раздражение которых вызывает определенный рефлекс, называют рецептивным полем рефлекса (например, участок кожи). Рецептивные поля разных рефлексов, находящиеся на поверхности кожи, могут накладываться одно на другое. Вследствие этого раздражение, наносимое на определенный участок кожи, в зависимости от его силы и состояния ЦНС может вызвать то один, то другой рефлекс.
Рефлексы отличаются большим многообразием. Их можно классифицировать по ряду признаков на разные группы. По происхождению рефлексы бывают условные, или приобретенные в процессе жизни, и безусловные, или врожденные, передающиеся от поколения к поколению.
В зависимости от места расположения рецепторов рефлексы подразделяют на экстерорецептивные, т. е. вызываемые раздражением рецептора на поверхности тела; интерорецептивные, или висцеральные, возникающие при раздражении рецепторов внутренних органов и сосудов; проприорецептивные, вызываемые раздражением рецепторов суставов, скелетных мышц, сухожилий.

|
Рис. 13.6. Схема рефлекторной дуги:
/ — афферентный нейрон; 2—афферентный (центростремительный) нерв; 3 — нервный центр; 4— эфферентный (центробежный) нерв; 5—эффектор (исполнительный орган)
Рефлексы подразделяют по характеру ответных реакций в зависимости от того, какие органы в ней участвуют, на моторные, или двигательные, в которых исполнительным органом служат мышцы; сосудодвигательные, проявляющиеся в сужении или расширении кровеносных сосудов; секреторные, которые заканчиваются секрецией желез.
Рефлексы классифицируют по месту расположения нервных центров на спиналъные, в которых принимают участие нейроны спинного мозга; бульварные, осуществляемые при участии нейронов продолговатого мозга; мезэнцефальные, осуществляемые при участии нейронов среднего мозга; диэнцефалъные, в которых участвуют нейроны промежуточного мозга; кортикальные, в осуществлении которых принимают участие нейроны коры больших полушарий головного мозга.
Рефлексы разделяют и по биологическому значению — на пищевые, оборонительные, половые, тонические, ориентировочные, локомоторные и др. По месту раздражения и ответной реакции рефлексы бывают кутано-висцеральные, осуществляемые с кожи на внутренние органы; висцеро-кутанные — с внутренних органов на кожу; висцеро-висцеральные — с внутренних органов на внутренние органы.
Рефлексы бывают моносинаптическими и полисинаптически-ми. Моносинаптические — это простейшие рефлексы, поступающие в ЦНС по афферентному пути, переключающиеся непосредственно на эфферентный нейрон. Но рефлекторные дуги большинства рефлексов состоят не из двух, а из большего количества нейронов: афферентный, один или несколько вставочных и эфферентный. Такие рефлекторные дуги называют многонейронными, или полисинаптическими. Наиболее простая из этих дуг образована тремя нейронами; в ней имеется два синапса.
Схемы моно- и даже полисинаптических рефлекторных дуг дают упрощенное представление о составе нейронов, участвующих в том или ином рефлекторном акте, без учета всей сложности путей распространения импульсов по многочисленным проводящим путям в ЦНС при всяком рефлексе.
По своему строению различают дуги вегетативного и соматического рефлексов. Различие состоит в том, что эфферентный путь вегетативного рефлекса состоит из двух нейронов — прегангли-онарного и постганглионарного нервного волокна, а их соединение осуществляется в вегетативных ганглиях (симпатических и парасимпатических).
В учении о рефлексе есть понятие обратной связи, или обратной афферентации. Согласно ему импульс от рецепторов органа-исполнителя (эффектора) повторно поступает в ЦНС для оценки и корректировки выполненной реакции. Этот поток импульсов называется вторичной афферентацией в отличие от первичной, которая вызвала этот рефлекс (рис. 13.7). По данному
 | |||
 |

|
Рис. 13.7. Схема рефлекторной дуги с обратной связью:
1 — рецептор; 2—афферентный нейрон; 3— промежуточный нейрон; 4— эфферентный нейрон; 5—эффектор (мышца); 6— проприорецептор (мышечное веретено); 7— нейрон обратной связи; стрелки — направление прохождения импульса
принципу осуществляются не только сложные поведенческие акты, но и поддерживается постоянство температуры тела, уровень сахара в крови, кровяное давление. Для примера можно привести следующий тест: перед испытуемым лежит молот, сделанный из картона, но имеющий вид настоящего молота. Он не знает, что молот легкий и, поднимая его, делает усилие. Так как масса предмета не соответствует данному усилию, получается некоординированный рывок. Но попросите испытуемого взять второй раз этот молот, и он возьмет его как легкий предмет, так как только что от рецепторов мышц его руки в ЦНС поступили сигналы о том, что предмет легкий.
Таким образом, принцип обратной связи обеспечивает такое совершенное управление процессами со стороны ЦНС, которого не может быть при односторонней связи.
СВОЙСТВА НЕРВНЫХ ЦЕНТРОВ
Нервным центром называется совокупность нейронов, необходимых для осуществления определенного рефлекса. С физиологической точки зрения нервный центр —это сложное сочетание нейронов, согласованно и координированно участвующих в регуляции функций и рефлекторной реакции. Нервные центры обладают рядом характерных свойств, определяемых особенностями синаптической передачи нервных импульсов и структурой нейронных цепей, образующих эти центры.
Односторонняя проводимость. В нервном волокне импульсы могут проводиться в обоих направлениях. В ЦНС возбуждение может распространяться только в одном направлении: с афферентного нейрона на эфферентный. Одностороннее проведение обусловлено тем, что передача возбуждения возможна через синапс только в одном направлении — от нервного окончания, секретирующего медиатор, к постсинаптической мембране. В обратном направлении возбуждающий постсинаптический потенциал не распространяется.
Задержка проведения возбуждения. По рефлекторной дуге проведение возбуждения происходит медленнее,
чем по нервному волокну. Это объясняется особенностями передачи возбуждения с одного нейрона на другой через синапс. При передаче импульса через синапс проходят следующие процессы:
выделение медиатора нервным окончанием в ответ на пришедший к нему по аксону импульс;
диффузия медиатора через синаптическую щель к постсинаптической мембране;
возникновение под влиянием медиатора возбуждающего пост-синаптического потенциала.
Задержка проведения возбуждения в синапсах является причиной того, что в нервных центрах импульсы распространяются медленнее, чем в нервных волокнах. Этим определяется время рефлекса, т. е. интервал от начала раздражения рецептора до появления ответной реакции. В течение этого времени происходят следующие процессы:
возбуждение рецепторов;
проведение возбуждения по центростремительным нервным волокнам к нервным центрам;
передача возбуждения с афферентных нейронов на эфферентные через один или несколько синапсов;
проведение возбуждения по центробежным нервным волокнам от ЦНС;
передача возбуждения с нерва на рабочий орган.
Таким образом, время рефлекса представляет собой сумму продолжительности всех перечисленных процессов. Это время называется центральной задержкой. Наиболее продолжительно время рефлекторных реакций внутренних органов, кровеносных сосудов и потовых желез, импульсы к которым передаются через вегетативную нервную систему.
Время рефлекса зависит от силы раздражения и от состояния ЦНС. Зная центральное время какого-либо рефлекса и учитывая, что на проведение через синапс необходимо 1,5...2 с-3, можно определить число синапсов, через которые проходит нервный импульс.
Иррадиация. Характер рефлекса в значительной степени зависит от интенсивности раздражения и числа активируемых рецепторов. Усиление раздражения приводит к расширению рецептивного поля рефлекса, в результате чего вовлекается большее число центральных нейронов. Этот процесс называется иррадиацией возбуждения и в значительной степени зависит от того, что отдельные афферентные нейроны вызывают подпороговую деполяризацию центральных нейронов.
Таким образом, иррадиация — это распространение возбуждения в виде широкой волны в ЦНС от центра к центру благодаря наличию большого количества коллатералей. Лучше всего иррадиация проявляется в ретикулярной формации: по ее восходящему отделу возбуждение диффузно распространяется в коре больших полушарий.
36 — 3389
 Суммирование возбуждений. Было открыто И. М. Сеченовым / в нервных центрах. Существует два вида суммирования: временное \ и пространственное. Суммирование происходит в результате возникновения или усиления рефлекса при действии слабых и частых раздражений, каждое из которых в отдельности или не вызывает ответную реакцию, или же эта реакция очень слабая.
Суммирование возбуждений. Было открыто И. М. Сеченовым / в нервных центрах. Существует два вида суммирования: временное \ и пространственное. Суммирование происходит в результате возникновения или усиления рефлекса при действии слабых и частых раздражений, каждое из которых в отдельности или не вызывает ответную реакцию, или же эта реакция очень слабая.
Если на лапку лягушки нанести подпороговое раздражение, то оно не вызывает ответной реакции, если же наносить несколько подпороговых раздражений через короткие интервалы времени, то лягушка сгибает ногу. Возникновение данного рефлекса обусловлено временным суммированием.
Если наносить одновременно подпороговое раздражение на два лежащих в пределах рецептивного поля участка кожи собаки, то происходит рефлекс почесывания. Каждое такое раздражение, действуя в отдельности, не вызывает рефлекса, при сочетании же возникает рефлекторная реакция. Таким образом, возникновение этого рефлекса обусловлено раздражением нескольких афферентных волокон. Такое явление получило название пространственного суммирования.
Трансформация ритма и силы — усиление или ослабление ритма или силы возбуждения, поступающего с периферии. В связи с этим частота импульсов, возникающих в нейроне, на котором заканчиваются синапсы другого нейрона, относительно независима от частоты импульсов, пришедших к этим синапсам. В ряде случаев в ответ на одиночный импульс, приложенный к афферентному нерву, мотонейрон по аксону посылает целую серию импульсов, следующих друг за другом с определенным интервалом. В нервных центрах может происходить и трансформация силы импульса, т. е. слабые импульсы усиливаются, а сильные ослабляются.
Утомление нервного центра проявляется в снижении и полном прекращении рефлекторного ответа при продолжительном раздражении афферентных нервных волокон. Утомление нервных центров связано с нарушением передачи возбуждения в межнейронных синапсах. Оно обусловлено резким уменьшением запасов медиатора в окончаниях аксонов, понижением чувствительности к медиатору постсинаптической мембраны, сокращением ее энергетических запасов. Но не все рефлекторные акты приводят к быстрому развитию утомления. К таким рефлексам можно отнести проприорецептивные тонические рефлексы.
Последействие — способность сохранять возбуждение в течение некоторого времени после прекращения действия раздражителя. Продолжительность последействия может во много раз превышать продолжительность раздражения, и обычно тем продолжительнее, чем сильнее раздражение и дольше оно действовало на рецептор. Существует два основных механизма, обусловливающих эффект последействия. Первый связан со следовой деполяризацией мембраны нейрона после длительного ритмичес-
кого раздражения. Однако следовая деполяризация может обусловить лишь кратковременный эффект последействия. Второй же механизм связан с циркуляцией нервных импульсов по замкнутым нейронным цепям. При таком включении нейронов возбуждение одного из них передается на другой, а по коллатерали их аксонов вновь возвращается к первой клетке.
Тонус — состояние незначительного возбуждения нервных центров. Редкие импульсы, непрерывно поступающие из нервных центров на периферию, обусловливают тонус скелетных мышц, гладких мышц кишечника, сосудов. В его поддержании участвуют как афферентные импульсы, поступающие непрерывно от периферических рецепторов в ЦНС, так и различные раздражители. Большую роль в этом процессе играет кольцевое взаимодействие между нервными центрами и периферией. В поддержании тонуса участвуют соответствующие центры продолговатого, среднего и промежуточного мозга.
Пластичность — свойство нервных центров изменять или перестраивать свою функцию. Например, П.К.Анохин (1935) сшивал у собаки центральный конец блуждающего нерва с периферическим концом лучевого. Волокна блуждающего нерва «врастали» в конечность, и на первых порах после операции сгибание или почесывание ее вызывало у собаки кашель и рвоту (функции, связанные с центром блуждающего нерва). Затем произошла функциональная перестройка центра вследствие свойственной ему пластичности, и конечность стала двигаться нормально. Большое значение в этих процессах играет кора больших полушарий, так как ее удаление делает невозможным проявление пластичности.
Восстановление нарушенной функции наблюдается также после разрушения или удаления отдельных участков ЦНС. Так, движения, резко нарушенные у животных в первое время после полного или частичного удаления мозжечка, удаления отдельных участков головного мозга или разрушения лабиринтов, через некоторое время в известной степени восстанавливаются вследствие пластичности нервных центров, но, как правило, этот процесс длительный.
|
|
|
