 |


Антибиотики, ингибирующие матричные биосинтезы
|
|
|
|
Тема 6. БИОСИНТЕЗ БЕЛКОВ И НУКЛЕИНОВЫХ КИСЛОТ. РЕГУЛЯЦИЯ БИОСИНТЕЗА БЕЛКА
Практическая значимость темы. Генетическая информация закодирована в последовательности нуклеотидов ДНК, находящейся в клеточном ядре. Сохранение, передача и реализация этой информации представляет собой сложный процесс, в котором участвуют многие вещества.
Генетическая информация, заключённая в каждой клетке организма, практически идентична. Реализация этой информации регулируется в ходе индивидуального развития организма и дифференцировки составляющих его клеток. Кроме того, чтобы организм мог приспосабливаться к изменяющимся условиям окружающей среды, система передачи генетической информации должна отвечать на внешние сигналы.
Ряд факторов, в том числе физические, химические и биологические воздействия, могут вызывать образование мутаций ДНК. Эти изменения передаются в процессе деления дочерним клеткам. Многие заболевания, в том числе опухолевые, являются следствием таких мутаций.
Понимание данных механизмов позволяет раскрыть причины наследственных заболеваний и наметить пути их коррекции, даёт возможность обоснованного применения фармакологических препаратов для лечения злокачественных новообразований и инфекционных болезней.
Цель занятия. После изучения данной темы студент должен знать особенности биосинтеза и роль нуклеиновых кислот в клетке, этапы биосинтеза белка и важнейшие механизмы регуляции этого процесса; уметь применять полученные знания для решения теоретических и практических задач.
Исходный уровень знаний.
1. Структурная организация белковых молекул.
2. Строение ДНК и РНК, их структурные и функциональные особенности, разновидности РНК.
|
|
|
3. Нуклеозидтрифосфаты, их роль в организме.
4. Механизмы регуляции активности ферментов.
Общая характеристика.
6.1.1. Матричный биосинтез – процесс сборки новых макромолекул из мономеров, последовательность которых запрограммирована с помощью нуклеиновых кислот. Молекулы, используемые в качестве программы в матричном биосинтезе, называют матрицами.
Тремя главными матричными биосинтезами, присущими всем без исключения живым организмам, являются репликация ДНК, транскрипция и трансляция.
· репликация ДНК происходит в ядре, предшествует делению клеток, в результате чего дочерние клетки получают полный набор генов;
· транскрипция также осуществляется в ядре, в ходе её образуются матричные, транспортные и рибосомальные РНК, участвующие в синтезе белка в клетке;
· трансляция происходит на рибосомах и приводит к образованию специфических клеточных белков.
Связь этих процессов отражена в основном постулате молекулярной биологии: направление потока информации от генотипа к фенотипу: ДНК → РНК → белок (стрелки обозначают направление передачи информации).
6.1.2. Кроме того, для некоторых видов вирусов характерны ещё два вида матричных синтезов:
· репликация РНК – синтез РНК на матрице РНК;
· обратная транскрипция – синтез ДНК с использованием в качестве матрицы молекулы РНК.
6.1.3. Попытаемся сформулировать общие закономерности, характерные для всех матричных биосинтезов.
1. Мономеры (нуклеотиды, аминокислоты) непосредственно в синтезе полимеров участвовать не могут; они должны находиться в активной форме – нуклеотиды – в виде нуклеозидтрифосфатов, аминокислоты – в виде соединений с тРНК.
2. Синтез всех полинуклеотидных и полипептидных цепей складывается из трёх основных этапов – инициации, элонгации и терминации.
3. На матрице имеется специальный сигнал или группа сигналов, позволяющие опознать кодирующий элемент, с которого начинается информация о синтезируемой цепи биополимера. Этот сигнал, как правило, не совпадает с точкой физического начала полимерной цепи матрицы. Инициация – процесс, в котором происходит присоединение первого мономерного звена к молекуле-матрице.
|
|
|
4. На каждый акт инициации биосинтеза приходится большое количество актов элонгации, т.е. соединения очередного мономера с растущей цепью. В элонгации участвуют 3 компонента: а) концевая группа синтезируемого полимера, б) кодирующий элемент матрицы, в) очередная молекула активного мономера. Все они должны быть зафиксированы определённым образом в активном центре фермента или рибосомы.
5. Каждый акт элонгации начинается с отбора субстратов путём перебора всех присутствующих субстратов в системе. Попадание в активный центр нужного субстрата является сигналом для осуществления ферментативной реакции соединения мономерного фрагмента с концом синтезируемой полимерной цепи. Присоединение мономера к растущей цепи служит сигналом для перемещения активного центра на один кодирующий элемент матрицы.
6. Конец продукта чаще всего не соответствует концу матрицы, на ней должен быть специальный сигнал, обеспечивающий прекращение роста цепи, т.е. терминацию.
7. Синтез биологически активной молекулы, как правило, не заканчивается терминацией. Образующийся полимер претерпевает ряд превращений, таких как частичный гидролиз и объединение нескольких цепей в одну, модификация мономеров в составе полимера, присоединение простетической части (к полипептиду) или апопротеина (к полинуклеотиду).
Репликация ДНК.
6.2.1. Репликация – процесс самоудвоения ДНК, или биосинтез дочерней молекулы ДНК, полностью идентичной исходной молекуле (матрице). Локализация процесса – клеточное ядро.
Условия, необходимые для репликации ДНК:
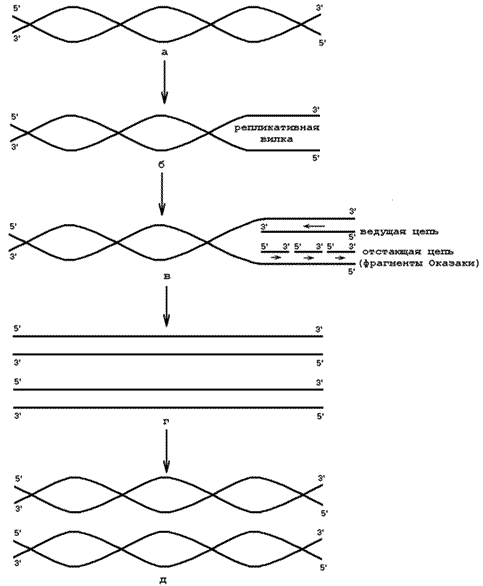
Рисунок 6.1. Репликация ДНК (объяснения в тексте).
1) Матрица – молекула ДНК (рисунок 6.1, а);
2) Расплетающие белки – разрывают водородные связи между комплементарными основаниями двойной спирали ДНК, в результате чего образуется репликативная вилка (рисунок 6.1, б);
3) Субстраты и источники энергии – дезоксирибонуклеозидтрифосфаты (дАТФ, дГТФ, дТТФ, дЦТФ). Присоединяются к азотистым основаниям полинуклеотидных цепей при помощи водородных связей по принципу комплементарности;
|
|
|
4) ДНК-полимераза – фермент, который формирует из нуклеозидтрифосфатов новые полинуклеотидные цепи за счёт образования 3’,5’-фосфодиэфирных связей. Синтез идёт в направлении от 5’-конца к 3’-концу. На одной ветви репликативной вилки синтезируется непрерывная цепь, на другой – фрагменты Оказаки (рисунок 6.1, в);
5) ДНК-лигаза – фермент, соединяющий фрагменты Оказаки в единую цепь (рисунок 6.1, г).
В результате образуются две идентичные молекулы ДНК (рисунок 6.1, д).
Транскрипция
6.3.1. Транскрипция – биосинтез РНК на матрице ДНК. Процесс транскрипции также происходит в клеточном ядре. Условия, необходимые для транскрипции:
1. Матрица – участок одной из цепей ДНК (рисунок 6.2, а);
2. ДНК-зависимая РНК-полимераза – главный фермент, участвующий в транскрипции. Место присоединения фермента к ДНК – промотор;
3. Субстраты и источники энергии – рибонуклеозидтрифосфаты (АТФ, ГТФ, УТФ, ЦТФ). Связываются с азотистыми основаниями транскрибируемой цепи ДНК водородными связями по принципу комплементарности. При участии ДНК-зависимой РНК-полимеразы образуются 3’,5’-фосфодиэфирные связи. Направление полимеризации цепи РНК – от 5’-конца к 3’-концу. Сигналом к окончанию транскрипции служат терминирующие последовательности нуклеотидов (рисунок 6.2,б).
В результате образуются первичные транскрипты – предшественники мРНК, тРНК и рРНК. Затем происходит их посттранскрипционное созревание (процессинг). Процессинг мРНК включает:
· вырезание неинформативных участков – интронов (рисунок 6.2, в);
· сращивание (сплайсинг) информативных участков – экзонов (рисунок 6.2, г);
· защиту концевых участков от действия нуклеаз (рисунок 6.2, д):
o присоединение 7-метилгуанилата («колпачок») к 5’-концу 5’,5’- фосфодиэфирной связью;
o присоединение полиаденилового нуклеотида к 3’-концу.
Образующаяся мРНК поступает из ядра в цитоплазму клеток.
|
|
|
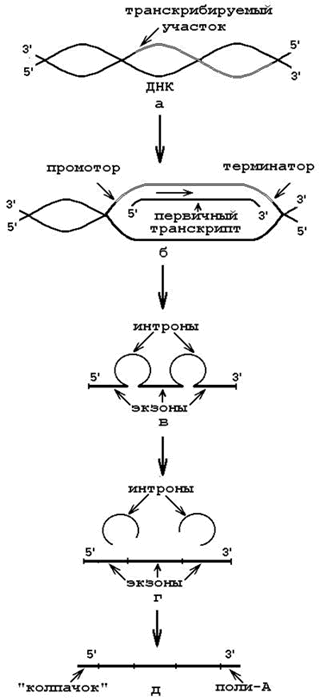
Рисунок 6.2. Транскрипция и созревание первичного транскрипта (объяснения в тексте).
Трансляция
6.4.1. Трансляция (от англ. translation – перевод) – перевод генетической информации, заключённой в мРНК, в линейную последовательность аминокислот в полипептидной цепи. Этот перевод осуществляется посредством генетического (биологического) кода.
6.4.2. Генетический код – последовательность нуклеотидов, соответствующая определённым аминокислотам. Генетический код характеризуется свойствами:
· код триплетный – каждой аминокислоте соответствует тройка (триплет) нуклеотидов – кодон. Всего существует 43 = 64 кодона. Из них 61 является смысловым (то есть кодирует определённую аминокислоту) и 3 – бессмысленными (терминирующими);
· код неперекрывающийся – один и тот же нуклеотид ДНК или РНК не может принадлежать одновременно двум соседним кодонам;
· код непрерывный – отсутствуют «знаки препинания», вставки между кодонами в полинуклеотидной цепи;
· код вырожденный (множественный) – некоторые аминокислоты могут кодироваться более, чем одним триплетом нуклеотидов (так как кодонов 61, а аминокислот – 20);
· код универсальный – смысл кодонов одинаков для организмов всех видов.
6.4.3. Аминокислоты и триплеты нуклеотидов, кодирующие их, не комплементарны друг другу. Поэтому должны существовать молекулы-адапторы, каждая из которых может взаимодействовать как с определённым кодоном, так и с соответствующей аминокислотой. Такими молекулами являются транспортные РНК (рисунок 6.3). Каждая тРНК содержит триплет нуклеотидов – антикодон, который комплементарен строго определённому кодону мРНК.
3’-конец тРНК (акцепторный участок) является местом присоединения аминокислоты, соответствующей кодону мРНК.
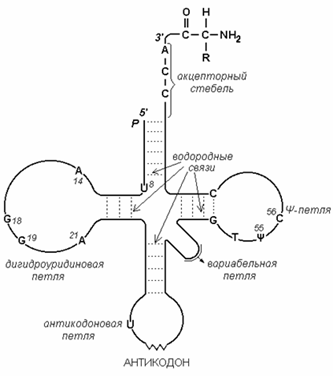
Рисунок 6.3. Схема строения транспортной РНК.
6.4.4. Активация аминокислот – подготовительный этап биосинтеза белка – включает связывание их со специфическими тРНК при участии фермента аминоацил-тРНК-синтетазы. Реакция происходит в цитоплазме клеток. Схема этой реакции показана на рисунке 6.4.

Рисунок 6.4. Реакция активации аминокислот.
6.4.5. Собственно процесс трансляции включает 3 стадии – инициации, элонгации, терминации и происходит на рибосомах.
Каждая рибосома состоит из большой и малой субчастиц (40S и 60S) и содержит аминоацильный (А) и пептидильный (П) участки. Пептидильный участок связывает инициирующую аминоацил-тРНК, все остальные аминоацил-тРНК присоединяются к аминоацильному участку.
Стадии трансляции представлены на рисунке.

Рисунок 6.5. Стадии трансляции (объяснения в тексте).
1) Стадия инициации – начало трансляции. Условия, необходимые для инициации:
|
|
|
· инициирующий кодон мРНК (АУГ);
· белковые факторы инициации;
· малая и большая субчастицы рибосомы;
· ГТФ (источник энергии для смыкания субчастиц рибосомы);
· ионы магния;
· инициирующая аминоацил-тРНК (метионил-тРНК) – связывается своим антикодоном с инициирующим кодоном мРНК в пептидильном участке рибосомы.
В результате образуется инициирующий комплекс: мРНК – рибосома – метионил-тРНК (рисунок 6.5, а).
2) Стадия элонгации – удлинение полипептидной цепи на 1 аминокислотный остаток – происходит в три шага:
· присоединение к инициирующему комплексу аминоацил-тРНК, соответствующей кодону, находящемуся в аминоацильном участке рибосомы (рисунок 6.5,б);
· транспептидация – образование пептидной связи между остатками аминокислот (рисунок 6.5, в). Источник энергии – ГТФ;
· транслокация – перемещение рибосомы относительно мРНК на 1 триплет (рисунок 6.5, г). Источник энергии – ГТФ. В ходе элонгации принимают участие белковые факторы.
Описанный процесс многократно повторяется (по количеству аминокислот в цепи).
3) Стадия терминации – окончание трансляции. Обеспечивается присутствием в цепи мРНК одного из терминирующих (бессмысленных) кодонов – УАА, УГА или УАГ. В освобождении полипептида участвуют белковые факторы терминации (рисунок 6.5, д). Когда в аминоацильном участке оказывается один из бессмысленных кодонов, факторы терминации стимулируют гидролазную активность пептидилтрансферазы. Благодаря этому гидролизуется связь между тРНК и пептидом. ГТФ для этой реакции не требуется. После этого пептидная цепь, тРНК и мРНК покидают рибосому, а её субчастицы диссоциируют.
Таким образом, трансляция мРНК приводит к формированию пептидной цепи со строго определённой последовательностью аминокислотных остатков. Следующий этап формирования белка – фолдинг, т.е. сворачивание пептидной цепи в правильную трёхмерную структуру. Если белок состоит из нескольких субъединиц, то фолдинг включает и объединение их в единую макромолекулу.
Считается, что небольшие белковые молекулы, содержащие около 100 аминоацильных остатков, могут самостоятельно принимать трёхмерную структуру, фолдинг более крупных полипептидных цепей требует участия специальных белков – шаперонов.
Посттрансляционные модификации белковой молекулы могут включать: частичный протеолиз; объединение нескольких полипептидных цепей в олигомерный белок; присоединение простетической группы; модификации аминокислотных остатков (гидроксилирование пролина в гидроксипролин в коллагене, метилирование аргинина в гистоне, йодирование тирозина в тироглобулине).
Ряд химических веществ, способных тормозить синтез белка, используется в лечении инфекционных заболеваний и злокачественных опухолей. К ним относятся многие антибиотики (см. таблицу 7).
Таблица 7
Антибиотики, ингибирующие матричные биосинтезы
| Антибиотики | Механизм действия |
| Противоопухолевые препараты:актиномицин Д, рубомицин С, митомицин С | Подавляют репликацию или транскрипцию, или оба эти процесса |
| Противобактериальные препараты: тетрациклин, левомицетин, эритромицин, стрептомицин | Ингибируют трансляцию в бактериальных (но не эукариотических) клетках |
Регуляция синтеза белка
6.5.1. В генетическом аппарате наряду со структурными генами, кодирующими структуру определённых белков, существуют участки ДНК, выполняющие регуляторные функции. Группа структурных генов, отвечающих за синтез ферментов одного метаболического пути, находится под контролем гена-оператора, расположенного рядом. Структурные гены в совокупности с геном-оператором составляют оперон, или транскриптон. Функция гена-оператора контролируется пространственно удалённым от него геном-регулятором, который продуцирует белок-репрессор, находящийся в активной либо в неактивной форме. Активный белок-репрессор способен связываться с геном-оператором и тормозить транскрипцию структурных генов, следовательно, подавлять синтез белков. Вещества, вызывающие инактивацию белка-репрессора, являются индукторами синтеза белка, оказывающие противоположный эффект – корепрессорами.
6.5.2. Существуют два механизма регуляции синтеза белка – индукция и репрессия. Примером оперона, который регулируется по механизму индукции, является лактозный оперон, в состав которого наряду с геном-оператором входят 3 структурных гена, кодирующие ферменты катаболизма лактозы (см. рисунок 6.6). Лактоза является индуктором данного оперона. При высокой концентрации лактозы в среде ферменты синтезируются, при низкой концентрации – нет.

Рисунок 6.6. Схема механизма индукции синтеза белка.
6.5.3. По механизму репрессии регулируется гистидиновый оперон, содержащий ген-оператор и 10 структурных генов, кодирующих ферменты, необходимые для биосинтеза гистидина (см. рисунок 6.7). Гистидин является корепрессором данного оперона. При высокой концентрации гистидина в среде синтез ферментов прекращается, при отсутствии гистидина они синтезируются.
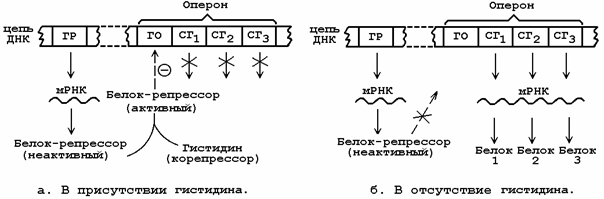
Рисунок 6.7. Схема механизма репрессии синтеза белка.
Контрольные вопросы:
1. Что такое матричные биосинтезы? Молекулы каких веществ выполняют роль матрицы и какие молекулы синтезируются при репликации, транскрипции, трансляции?
2. Что такое репликация ДНК? Какие условия необходимы для её протекания?
3. Дайте определение понятия транскрипции, укажите её важнейший фермент и его локализацию в клетке.
4. Укажите особенности процесса транскрипции (матрица, субстраты, источники энергии, фермент, направление полимеризации цепи).
5. Что понимают под интронами и экзонами? Опишите процессинг при образовании мРНК.
6. Дайте определение понятия процессинга мРНК и укажите, какие операции он включает.
7. Изобразите схематично структуру транспортной РНК. Укажите её важнейшие функциональные участки и их назначение.
8. Изобразите схематично структуру рибосомы. Укажите её важнейшие функциональные участки и их назначение при трансляции.
9. Дайте определение понятия генетического кода и охарактеризуйте его основные свойства.
10. Напишите схему реакции аминокислот в процессе биосинтеза белка, назовите ферменты.
11. Дайте определение понятия «трансляция». Назовите основные стадии этого процесса и их назначение, необходимые компоненты и условия.
12. Представьте в виде схемы инициирующий комплекс белоксинтезирующей системы. Перечислите условия, необходимые для инициации синтеза белка.
13. Дайте определение понятия элонгации. Представьте в виде схемы процесс элонгации на рибосоме, укажите необходимые условия, ферменты, факторы.
14. Опишите процесс терминации синтеза белка, укажите условия и факторы, способствующие терминации.
15. Представьте в виде схемы процесс терминации биосинтеза белка. Перечислите варианты посттрансляционной модификации молекул белков.
16. Какие соединения являются донорами энергии для биосинтеза белка из аминокислот? Укажите затраты энергии на каждом этапе биосинтеза белка.
17. Приведите примеры и укажите механизм действия ингибиторов биосинтеза белка, используемых в качестве лекарственных средств.
18. Дайте определение понятия «оперон». Опишите процесс регуляции биосинтеза белка путём индукции.
19. Представьте в виде схемы регуляцию синтеза белка на уровне транскрипции. Назовите регуляторные факторы и опишите механизм регуляции путём репрессии.
|
|
|
