 |


Рефлекторная дуга вегетативной
|
|
|
|
Нервной системы
В простой трехнейронной вегетативной рефлекторной дуге (рис.2), как и в соматической, выделяют те же три звена, а именно: рецепторное, образованное чувствительным (афферентным) нейроном, ассоциативное, представленное вставочным (ассоциативным) нейроном и эффекторное звено, образованное двигательным (эффекторным) нейроном, передающим возбуждение на тот или иной рабочий орган. Нейроны связаны между собой синапсами, в которых с помощью медиаторов происходит передача нервного импульса с одного нейрона на другой.
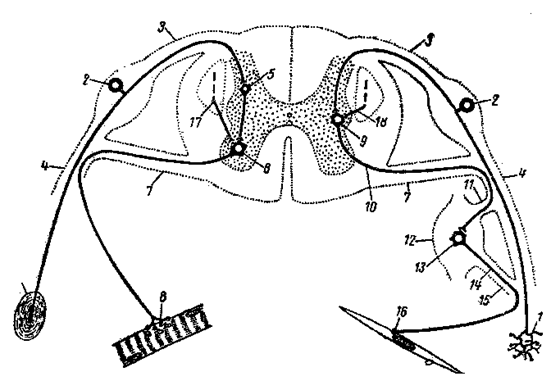
Рис. 2. Схема рефлекторных дуг соматического (слева) и вегетативного (справа) типов, замыкающихся в спинном мозге.
1- рецептор; 2- чувствительный нейрон спинномозгового ганглия; 3- дорсальный корешок; 4- спинномозговой нерв; 5- вставочный нейрон; 6- двигательный нейрон переднего рога; 7- вентральный корешок; 8- двигательное нервное окончание скелетной мышцы; 9- нейрон симпатического ядра бокового рога; 10- преганглионарное волокно; 11- белая соединительная ветвь; 12- периферический вегетативный ганглий; 13- эффекторный нейрон; 14- постганглионарное волокно; 15- серая соединительная ветвь; 16- двигательное нервное окончание на гладкой мышце; 17и 18- волокна пирамидного пути.
Чувствительные нейроны представлены псевдоуниполярными клетками спинномозгового узла, так же как и в соматической нервной системе. Их периферические отростки заканчиваются рецепторами в органах. Поэтому информация о состоянии органов растительной и животной жизни стекается в спинномозговые узлы, и в этом смысле они являются смешанными соматически-вегетативными узлами. Центральный отросток чувствительного нейрона в составе заднего корешка вступает в спинной мозг и нервный импульс переключается на вставочный нейрон, клеточное тело которого расположено в боковых рогах (латерально-промежуточное ядро тораколюмбального или сакрального отделов) серого вещества спинного мозга.
|
|
|
Вставочный нейрон отдает аксон, который покидает спинной мозг в составе передних корешков и достигает одного из вегетативных узлов, где вступает в контакт с эффекторным (двигательным) нейроном.
Таким образом, второе звено вегетативной рефлекторной дуги отличается от соматической, во-первых, местом локализации тела вставочного нейрона, во-вторых, протяженностью и положением аксона, который в отличие от соматической нервной системы никогда не остается в пределах спинного мозга. Еще большие различия в строении третьего звена рефлекторной дуги. В отличие от соматической рефлекторной дуги, где двигательные нейроны расположены в передних рогах спинного мозга, для вегетативной рефлекторной дуги характерно расположение двигательного нейрона за пределами центральной нервной системы - в вегетативных узлах, аксоны которых направляются к рабочему органу, а это значит, что весь эфферентный путь подразделяется на два участка: предузловой (преганглионарный) - аксон вставочного нейрона и послеузловой (постганглионарный) - аксон двигательного нейрона вегетативного узла. Таким образом, в вегетативной рефлекторной дуге эфферентный периферический путь двухнейронный.
В простой трехнейронной вегетативной рефлекторной дуге, замыкающейся в пределах мозгового ствола, тело первого нейрона располагается в чувствительных узлах черепных нервов, второго - в вегетативных ядрах черепных нервов (мезенцефалический и бульбарный отделы) и третьего - в вегетативных узлах.
Достигая эффекторов (гладких мышц и желез), нервные импульсы вызывают сокращение мышцы или изменение секреторной деятельности железы, что в свою очередь вызывает раздражение рецепторов этих органов и отсюда поток импульсов по афферентным волокнам направляется обратно в ядра спинного или головного мозга, неся ежемоментную информацию о состоянии данного органа. Наличие обратной связи (обратной афферентации), с одной стороны позволяет осуществлять контроль за правильностью исполнения команд, с другой - вносить дополнительную своевременную коррекцию в выполнении ответной реакции организма.
|
|
|
Таким образом, в основе строения и функции вегетативной нервной системы, как и соматической, лежит замкнутая кольцевая цепь рефлексов, которая способствует наиболее полному приспособлению организма к окружающей среде.
ВЕГЕТАТИВНЫЕ УЗЛЫ
Узлы вегетативной нервной системы устроены однотипно, но отличаются друг от друга своей локализацией. По этому топографическому признаку их делят условно на три группы (порядка). Узлы I порядка, околопозвоночные (паравертебральные), ganglia trunci sympathici, образуют симпатический ствол, расположенный по сторонам позвоночного столба. Узлы II порядка, предпозвоночные (превертебральные) или промежуточные, ganglia intermedia, расположены впереди позвоночника, входят в состав вегетативных сплетений и также как и узлы I порядка относятся к симпатическому отделу вегетативной нервной системы. Узлы III порядка составляют конечные (терминальные) узлы, ganglia terminalia. Они в свою очередь разделяются на околоорганные и внутриорганные и относятся к парасимпатическим узлам.
Различная степень удаленности вегетативных узлов от спинного мозга или мозгового ствола, с одной стороны, и рабочего органа, с другой, сказывается на длине преганглионарных и постганглионарных волокон. Наибольшую длину имеют преганглионарные волокна, связанные с узлами III порядка. Постганглионарные волокна этих узлов, наоборот, очень короткие. Обратная картина наблюдается при рассмотрении волокон, связанных с узлами I порядка. Здесь преганглионарные волокна короткие, а постганглионарные - длинные. Преганглионарные волокна покрыты миелиновой оболочкой, благодаря чему имеют белый цвет. Их диаметр чаще составляет 2-3,5 мкм, а скорость распространения импульсов колеблется от 3 до 18 м/сек. Постганглионарные волокна лишены миелина, серого цвета, диаметр - до 2 мкм, скорость проведения импульса - 1-3 м/сек.
|
|
|
Снаружи вегетативный узел покрыт соединительнотканной капсулой, от которой внутрь отходят тяжи, между которыми располагаются группы клеток. Тело каждого нейрона имеет тонкую соединительнотканную капсулу. Между ней и телом нейрона - клетки сателлиты, которые выполняют опорную, защитную и трофическую функции.
В узлах выделяют три типа нейронов:
1. Клетки Догеля первого типа - мультиполярные с множеством коротких ветвящихся дендритов и одним длинным аксоном, направляющимся к рабочему органу (рис. 3). Это двигательные нейроны.
2. Клетки Догеля второго типа - чувствительные, мультиполярные с длинными дендритами, которые покидают узел и заканчиваются рецепторами на гладкомышечных или железистых клетках. Аксон может заканчиваться здесь же в ганглии, или направляться в соседние.
3. Клетки Догеля третьего типа представляют местные ассоциативные элементы, соединяющие своими отростками несколько клеток I и II типов как своего узла, так и соседних узлов.

| Рис.3. Различные типы нейронов соматической (а) и вегетативной (б) частей нервной системы (из А.Г.Кнорре и И.Д.Лев). 1- пирамидная клетка коры головного мозга; 2- клетка Пуркинье из коры мозжечка; 3- клетка-зерно из коры мозжечка; 4- двигательная клетка переднего рога спинного мозга; 5- биполярная клетка сетчатки; 6- униполярная клетка сетчатки; 7- клетка 1 типа Догеля; 8- клетка II типа Догеля; 9- клетка промежуточно-латерального ядра бокового рога спинного мозга. |
Вегетативные узлы выполняют следующие функции:
1. Передачу нервного импульса с преганглионарной на постганглионарную часть эфферентного пути. Эта функция обусловлена наличием в узле двигательных нейронов.
2. Рефлекторную или замыкательную. Благодаря наличию не только двигательных, но и чувствительных нейронов в вегетативных узлах могут замыкаться периферические рефлекторные дуги, что превращает их в периферические нервные центры, способные осуществлять местные рефлексы на основе принципа саморегуляции.
3. Рецепторную, так как в узлах имеются рецепторы, которыми заканчиваются дендриты чувствительных нейронов спинномозговых и черепных узлов. Благодаря этим центростремительным связям состояние и деятельность вегетативных узлов находятся под контролем центральной нервной системы.
4. Через вегетативный узел проходят транзитно афферентные и эфферентные волокна.
Кроме анатомически обособленных ганглиев, по ходу вегетативных ветвей периферических нервов встречается большое количество нервных клеток, мигрировавших сюда в ходе эмбрионального развития.
|
|
|
