 |


Низшие растения, или Водоросли
|
|
|
|
Водоросли
Систематика растений занимается изучением и описанием видов растений и распределением их по группам на основе сходства строения и родственных связей между ними, созданием классификации.
Основными таксономическими категориями при классификации растений являются вид, род, семейство, порядок, класс, отдел, царство. Иногда используют промежуточные таксономические категории: подвид, надрод, подцарство, надцарство и другие.
Таблица 1. Таксономические категории и таксоны на примере картофеля:
| Таксономические категории | Таксоны |
| Царство | Растения |
| Отдел | Покрытосеменные |
| Класс | Двудольные |
| Порядок | Пасленовые |
| Семейство | Пасленовые |
| Род | Паслен |
| Вид | Картофель, или паслен клубненосный |
Низшие растения, или Водоросли
Общая характеристика. Водоросли – большая сборная группа фотосинтезирующих, преимущественно водных, фотоавтотрофных эукариотических растений. Для большинства водорослей характерно: в основном водная среда обитания, но большое число видов встречается и на суше (на поверхности почвы, влажных камнях, коре деревьев и т.д.).
Большинство водорослей находится в толще воды во взвешенном состоянии или активно плавает (фитопланктон), некоторые ведут прикрепленный образ жизни (фитобентос). Зеленые водоросли обитают в прибрежной зоне на небольшой глубине, бурые содержат пигменты, позволяющие им жить на глубине до 50 м, а набор фотосинтетических пигментов красных водорослей позволяет им обитать на глубине100-200 м, а отдельные представители обнаружены на глубине до 500 м.
Тело водорослей может быть одноклеточным, колониальным или многоклеточным. Если это многоклеточный организм, то его тело не дифференцировано на органы и ткани и называется таллом, или слоевище. У сложно организованных водорослей может наблюдаться элементарная дифференцировка тела, имитирующая органы высших растений – появляются ризоиды, стеблевидные и листовидные образования.
|
|
|
Строение клеток. Клетки большинства водорослей имеют клеточную стенку, образованную целлюлозой и пектином (только у примитивных подвижных одноклеточных и колониальных водорослей, у зооспор и гамет клетки ограничены лишь плазмалеммой), клеточная стенка почти всегда покрыта слизью. Протопласт клеток состоит из цитоплазмы, одного или нескольких ядер и хроматофоров (пластид), содержащих хлорофилл и другие пигменты; в хроматофорах имеются особые образования – пиреноиды – белковые тельца, вокруг которых накапливается крахмал, образующийся в процессе фотосинтеза. Вакуоли, как правило, хорошо развиты; иногда (особенно в подвижных клетках) имеются особые сократительные вакуоли; большинство подвижных водорослей имеют жгутики и светочувствительное образование – глазок, или стигму, благодаря которому водоросли обладают фототаксисом (способностью к активному движению всего организма по направлению к свету).
Питание автотрофное, но имеются виды сапрофиты и паразиты.
Размножение бесполое и половое, бесполое размножение осуществляется с помощью зооспор (подвижных) или спор (неподвижных). Бесполое размножение также может осуществляться с помощью вегетативного размножения путем фрагментации таллома, деления клеток одноклеточных водорослей, у колониальных водорослей – за счет распада колоний.
Половое размножение происходит путем образования множества специализированных половых клеток – гамет и их слияния (оплодотворения), что представляет собой половой процесс. В результате слияния образуется зигота, которая покрывается толстой защитной оболочкой. После периода покоя (реже сразу же) зигота прорастает в новую особь, образующуюся в основном путем мейотического деления (зиготическая редукция). Этим завершается половое размножение. Формы полового процесса водорослей – изогамия, гетерогамия, оогамия. Для некоторых водорослей половой процесс осуществляется в форме конъюгации. У высокоорганизованных водорослей гаметы развиваются в специальных органах полового размножения: яйцеклетки – в оогониях, сперматозоиды – в антеридиях. У диплоидных водорослей происходит спорическая редукция – мейоз происходит при образовании зооспор, из которых развиваются гаметофиты. Гаметофиты митотически образуют гаметы, и при образовании зиготы образуется диплоидная водоросль, формирующая споры – спорофит. У гаплоидных водорослей споры и гаметы могут развиваться в клетках одной и той же особи, митотически.
|
|
|
Происхождение водорослей обычно связывают с поглощением эукариотической аэробной клеткой цианобактерий, которые стали хлоропластами. В последнее время появилась гипотеза, которая дает объяснение, почему у различных отделов водорослей хлоропласты резко отличаются по строению и набору фотосинтетических пигментов – предками хлоропластов у разных групп водорослей, возможно, были различныефотосинтезирующие бактерии. С другой стороны, одни и те же фотосинтезирующие бактерии могли захватываться разными организмами-носителями, находящимися на разных стадиях эволюционного развития. А сходство митохондрий у всех эукариотических организмов свидетельствует о том, что митохондрии произошли от единого предка, наиболее близкого к современной аэробной несерной пурпурной бактерии и симбиоз анаэробного гетеротрофного прокариота и бактерии-окислителя сложился раньше, чем в состав симбиотического организма вошли фотосинтезирующие бактерии.

|
| Рис.. Многообразие растений |
Многообразие. Обычно водоросли подразделяют на несколько отделов: красные (выделяемые в самостоятельное подцарство Багрянки), бурые, зеленые, золотистые, желто-зеленые, диатомовые, харовые и эвгленовые водоросли (образуют подцарство Настоящие водоросли).

|
| Рис. 58. Формы таллома багрянок. |
Красные водоросли, или багрянки. Одно из подцарств царства Растения. Среди багрянок встречаются какодноклеточные, так и многоклеточные нитчатые и пластинчатые водоросли (рис.). Из 4000 видов только 200 приспособились к жизни в пресных водоемах и на почве, остальные – обитатели морей. Окраска красных водорослей разнообразна, она определяется различным количественным содержанием пигментов: хлорофиллы а и d, каротиноиды и фикобиллины: красный (фикоэритрин) и синий (фикоцианин). Причем окраска водорослей различна на разной глубине, на мелководье они желто-зеленые, затем розовые и на глубине более 50 м становятся красными. Максимальная глубина, на которой находили багрянки – 500 м, где они используют сине-фиолетовые длины волн солнечного света. Чем короче длина волны, тем больше ее энергия, поэтому на самую большую глубину проникают световые волны с наиболее короткой длиной волны. Причем водолазам они кажутся черными, настолько эффективно они поглощают весь падающий на них свет, красными они выглядят на поверхности. Пигменты сосредоточены в хроматофорах, имеющих вид зерен или пластинок, пиреноидов нет.
|
|
|
Клеточная стенка – пектиново-целлюлозная, способная к сильному ослизнению, в результате чего у некоторых водорослей весь таллом приобретает слизистую консистенцию. В стенках у многих может откладываться углекислый кальций (СаСО3) или магний (MgCO3).
Продуктом ассимиляции является багрянковый крахмал, по строению близкий к гликогену. В отличие от обычного крахмала при окрашивании йодом он приобретает буро-красный цвет.
Наблюдается чередование полового (гаплоидного,n) и бесполого (диплоидного 2n) поколений. Бесполое размножение красных водорослей осуществляется с помощью гаплоидных безжгутиковых спор, развивающихся мейотически в спорангиях (спорическая редукция). Для багрянок характерен оогамный половой процесс.
Из спор развиваются гаметофиты, на гаметофитах образуются безжгутиковые мужские и женские гаметы. Большинство багрянок – двудомные растения. Из зиготы развивается диплоидный спорофит. Гаметофит и спорофит по внешнему виду неразличимы. Вегетативное размножение частями таллома характерно лишь для низкоорганизованных багрянок.
|
|
|
Отсутствие жгутиковых форм – характерный признак багрянок. Предполагают, что багрянки произошли от эукариот, еще не имеющих жгутиков, на этом основании их выделяют в отдельное подцарство.
Багрянки имеют большое практическое значение. Из них получают агар-агар, использующийся в кондитерской и микробиологической промышленности, многие из них являются сырьем для получения клея. Из золы багрянок получают йод и бром. Некоторые красные водоросли используются на корм скоту. В Японии, Китае, на островах Океании и в США багрянки используются в пищу. Порфира считается деликатесом. Красная водоросль хондрус используется для получения каррагенов – особых полисахаридов, подавляющих размножение вируса СПИДа.
ОтделБурые водоросли. Отдел включает около 1500 видов многоклеточных, преимущественно макроскопических (до 60-100 м) водорослей, ведущих прикрепленный (бентосный) образ жизни. Чаще всего они встречаются в прибрежных мелководьях всех морей и океанов, иногда вдали от берега (например, в Саргассовом море).
Строение. Талломы бурых водорослей имеют наиболее сложное строение среди водорослей. Одноклеточные и колониальные формы отсутствуют. У высокоорганизованных клетки таллома отчасти дифференцируется, образуя тканеподобные анатомические структуры (например, ситовидные трубки с косыми перегородками). В результате этого происходит образование “стеблевой” и “листовой” частей таллома, выполняющих неоднородные функции. В субстрате водоросли закрепляются с помощью ризоидов.
Клетки бурых водорослей одноядерные с многочисленными хроматофорами, имеющими вид дисков или зерен. Бурая окраска водорослей обусловлена смесью пигментов (хлорофилла, каротиноидов, фукоксантина). Основным запасным веществом является ламинарин (полисахарид с иными, чем у крахмала, связями между остатками глюкозы), откладывающийся в цитоплазме. Клеточные стенки сильно ослизняются. Слизь помогает удерживать воду и тем самым препятствует обезвоживанию, что важно для водорослей приливно-отливной зоны.
Размножение половое и бесполое. Формы полового процесса различны – изогамия, гетерогамия, оогамия. Происходит чередование бесполого поколения – спорофита и полового – гаметофита. Спорофит и гаметофит по размеру и форме могут быть как одинаковыми, так и различными. Бесполое размножение происходит с помощью многочисленных двужгутиковых зооспор, мейотически образующиеся в одноклеточных, реже многоклеточных зооспорангиях спорофита. На гаметофитах (n) образуются оогонии и антеридии, в которых образуются гаметы, из зиготы развивается спорофит. Вегетативное размножение осуществляется частями таллома.
|
|
|
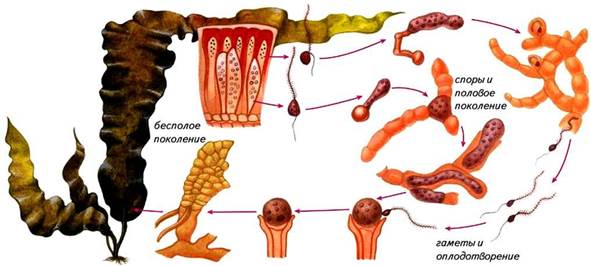
Ламинария. Представители рода ламинария известны под названием «морская капуста» (рис.). Они широко распространены в северных морях. Зрелый спорофит ламинарии диплоидное растение длиной от 0,5 до 6 и более метров.
|
| Рис.. Чередование поколений у ламинарии |
Слоевище ламинарии имеет одну или несколько листоподобных пластинок, располагающихся на простом или разветвленном стеблевидном образовании, прикрепленном к субстрату ризоидами. Стеблевидное образование с ризоидами многолетнее, а пластинка ежегодно отмирает и весной вновь отрастает.
Размножение. На поверхности пластинок формируются зооспорангии, в которых в результате мейотического деления образуются гаплоидные зооспоры с двумя неравными жгутиками. Они прорастают в микроскопические нитчатые гаметофиты, на которых образуются половые органы. Половой процесс оогамный. В оогониях и антеридиях образуется по одной гамете, после оплодотворения из зиготы без периода покоя развивается диплоидный спорофит. Таким образом, у ламинарии наблюдается чередование поколений, диплоидный спорофит образует зооспоры из которых развиваются гаплоидные гаметофиты.
Типичными представителями бурых водорослей является ламинария, макроцистис (его громадное слоевище достигает в длину 50-60 м), фукус, саргассум.
Значение. Будучи автотрофами, водоросли являются основными продуцентами (т. е. производителями) органических веществ в различных водоемах. Кроме того, в процессе фотосинтеза они выделяют кислород, создавая тем самым благоприятные условия для жизни не только водных, но и наземных организмов.
Водоросли играют огромную роль в жизни человека: являются кормом для многих промысловых рыб и других животных, служат добавками в различных питательных смесях, входят в состав комбикормов, некоторые водоросли (например, «морскую капусту») употребляют в пищу. Клетки бурых водорослей поверх целлюлозной клеточной стенки порыты пектином, состоящим из альгиновой кислоты или ее солей, при смешивании с водой (в соотношении 1/300) альгинаты образуют вязкий раствор. Альгинаты используются в пищевой промышленности (для получения пастилы, мармеладов), в парфюмерии (изготовление гелей), в медицине (для изготовления мазей), в химической промышленности (для изготовления клеев, лаков). В текстильной промышленности с их помощью делают невыцветающие и непромокаемые ткани. Морские водоросли используются для получения удобрений, йода, брома. Йод получали раньше исключительно из бурых водорослей. Бурые водоросли могут служить в качестве индикатора местонахождения золота, они способны накапливать его в клетках слоевища.
Отдел Зеленые водоросли. Отдел объединяет около 13000 видов, это самый обширный отдел среди водорослей. Отличительная особенность – чисто зеленый цвет слоевищ, вызванный преобладанием хлорофилла над другими пигментами. Распространены повсеместно. В основном зеленые водоросли обитатели пресных водоемов, но есть и морские виды. Некоторые обитают на суше. Есть виды, вступающие в симбиотические отношения с некоторыми животными (губками, кишечнополостными, оболочниками) и грибами.
Строение. Зеленые водоросли представлены одноклеточными, колониальными и многоклеточными формами. Клетки имеют плотную целлюлозно-пектиновую оболочку, бывают одноядерные или многоядерные. В цитоплазме находятся хроматофоры с пигментами (в основном хлорофилл a и b,). Кроме хлорофилла, в клетках содержатся каротиноиды, ксантофиллы и другие пигменты. Хлоропласты сходны с пластидами высших растений. Основным запасным веществом, накапливающимся в хлоропластах, является крахмал.
Размножение. Большинство зеленых водорослей гаплоидны. Редукция генетического материала происходит после образования зиготы – зиготическая редукция. Но часто наблюдается и правильное чередование полового и бесполого поколений.
Зеленые водоросли считаются предками наземных растений: они имеют одинаковые наборы фотосинтетических пигментов, оболочка содержит не только целлюлозу, но и пектин, запасное вещество – крахмал, накапливаются запасные питательные вещества не в цитоплазме (как у других водорослей), а в пластидах.

|
| Рис. 60. Строение хламидомонады: 1 – две пульсирующие вакуоли; 2 – стигма; 3 – цитоплазма; 4 – ядрышко; 5 – ядро; 6 – хроматофор; 7 – жгутик; 8 – пиреноид; 9 – зерна крахмала; 10 – клеточная стенка. |
РодХламидомонада. В переводе – единичный организм, покрытый древнегреческой одеждой – хламидой. Одноклеточные водоросли, обитающие преимущественно в мелких водоемах, загрязненных органическими веществами (рис. 60). Клетка хламидомонады имеет округлую или овальную форму, передний конец заострен в виде носика. На нем располагаются два одинаковой величины жгутика, с помощью которых хламидомонада передвигается в воде. Оболочка клетки пектиново-целлюлозная. В центре клетки располагается чашевидный хроматофор с крупным пиреноидом. В углублении хроматофора располагается ядро. На переднем конце клетки находятся стигма и пульсирующие вакуоли.
Размножается хламидомонада как бесполым, так и половым путем. В жизненном цикле преобладает гаплоидная фаза. При бесполом размножении хламидомонада теряет жгутики, содержимое клетки дважды делится митотически, и под оболочкой материнской клетки образуются четыре дочерние. Каждая из них выделяет оболочку и образует жгутики, превращаясь в зооспоры.
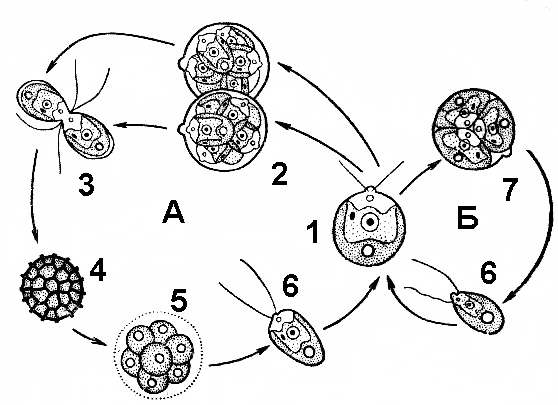
|
| Рис. 61. Жизненный цикл хламидомонады: А — половое размножение (1 — материнское растение; 2 — образование гамет; 3 — копуляция гамет; 4 — зигота; 5 — прорастание зиготы; 6 — зооспора); Б — бесполое размножение (7 — образование зооспор). |
Под воздействием ферментов оболочка материнской клетки разрушается, и они выходят наружу, растут до размеров материнской и тоже переходят к бесполому размножению (рис. 61).
Половой процесс у многих видов хламидомонады происходит по типу изогамии. Содержимое клетки делится, образуя от 8 до 32 гамет, которые напоминают зооспоры, но имеют более мелкие размеры. Клетки с разным половым знаком сливаются. Образовавшаяся зигота покрывается толстой оболочкой и впадает в период покоя. При наступлении благоприятных условий содержимое зигоспоры делится мейотически, и образуются четыре гаплоидные клетки, каждая из которых становится новой хламидомонадой.
У некоторых видов половой процесс осуществляется по типу гетерогамии (обе гаметы подвижны, но женская крупнее мужской) или по типу оогамии (женская гамета неподвижна).
Род Хлорелла. Одноклеточная водоросль, обитающая в пресных и соленых водоемах, на влажной почве, скалах (рис. 62). Клетки имеют вид зеленых шариков диаметром до 15 мкм. Жгутиков, глазков и сократительных вакуолей не имеет. В клетках имеется чашевидный хроматофор с пиреноидом или без него и мелкое ядро. Хлорелла гораздо более эффективно использует солнечную энергию для фотосинтеза. Если наземные растения используют около 1% солнечной энергии, то хлорелла – 10%. Половой процесс для этой водоросли не известен. Бесполое размножение происходит путем митотического деления содержимого материнской клетки дважды или трижды. В результате деления формируется четыре или восемь неподвижные споры (апланоспоры). После разрыва материнской оболочки клетки выходят наружу, увеличиваются в размерах и делятся вновь.

|
| Рис.. Размножение хлореллы |
Хлорелла интересна тем, что ее клетки содержат большое количество питательных веществ – 50 полноценных белков, жирные масла, углеводы, витамины А, В, С и К и даже антибиотики (причем витамина С в ней в 2 раза больше, чем в соке лимона). Она размножается так интенсивно, что за сутки происходит тысячекратное увеличение числа ее клеток.
Хлорелла стала первой водорослью, которую человек стал выращивать в культуре. Она использовалась в качестве экспериментального объекта для изучения некоторых этапов фотосинтеза. В некоторых странах (США, Япония, Израиль) созданы опытные установки для выращивания хлореллы и изучалась возможность использования хлореллы как источника питания для человека. Японцы научились перерабатывать хлореллу в белый порошок, богатый белками и витаминами. Его можно добавлять в муку для выпечки хлебобулочных изделий. Кроме того, хлорелла используется как источник дешевых кормов для скота и при биологической очистке сточных вод.
Класс Улотриксовые. Многоклеточные водоросли, слоевище которых нитевидное или пластинчатое. Наиболее известные представители относятся к роду Улотрикс и роду Ульва. Неветвящиеся нити улотрикса, прикрепляясь к подводным предметам – камням, сваям, корягам и т.д., образуют зеленые дерновинки. Все клетки (за исключением вытянутой в длину бесцветной ризоидальной клетки, с помощью которой происходит прикрепление водоросли) имеют сходное строение. В центре клетки находится ядро и хроматофор, имеющий форму незамкнутого кольца. В хроматофоре находится несколько пиреноидов. Рост нити в длину происходит за счет деления клеток в поперечном направлении. Произрастает в быстротекущих реках, ведет прикрепленный образ жизни (рис. 65).
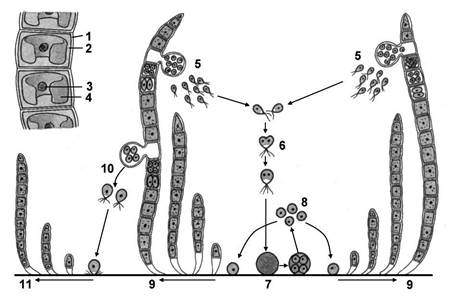
|
| Рис. 65. Улотрикс: 1 – клеточная оболочка; 2 – цитоплазма; 3 – ядро; 4 – хроматофор; 5 – образование гамет; 6 – копуляция гамет; 7 – зигота; 8 – мейоз и образование четвырех гаплоидных клеток; 9 – развитие улотрикса; 10 – образование четырехжгутиковых зооспор; 11 – развитие улотрикса из зооспор. |
При благоприятных условиях улотрикс размножается зооспорами, имеющими по четыре жгутика. Они образуются в четном количестве (2, 4, 8 и более). Зооспоры бывают разных размеров – крупные и мелкие. Способность к активному перемещению зооспор способствует расселению улотрикса.
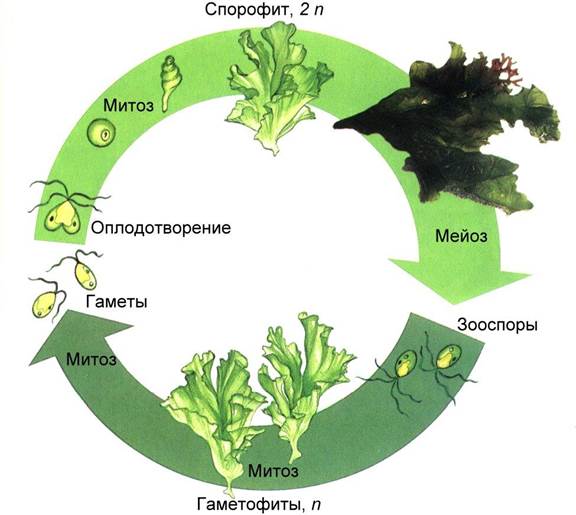
|
| Рис.. Чередование поколений у ульвы |
Половой процесс происходит по типу изогамии. Отдельные клетки нити превращаются в гаметангии, в которых образуются двужгутиковые гаметы. При слиянии гамет образуется четырехжгутиковая зигота. Затем она отбрасывает жгутики и переходит в состояние покоя.
В дальнейшем зигота редукционно делится, давая начало четырем клеткам, каждая из которых образуется новую нить.
Важная эволюционная линия связана с переходом от нитчатого слоевища к пластинчатому. Именно такая форма слоевища у представителей рода Ульва (морской салат). Внешне ульва напоминает тонкий зеленый лист целлофана, ее слоевище до 150 см состоит из двух слоев клеток. Для ульвы характерно чередование поколений, причем диплоидный спорофит и гаплоидные гаметофиты внешне не отличаются. Такое чередование поколений называется изоморфным.
Род Спирогира. Зеленые нитчатые водоросли длиной до 8-10 см (рис. 63). Многочисленные виды спирогир обитают в пресных водоемах, в стоячей воде. Скопления нитей спирогиры образуют тину. Нити неветвящиеся, образованные одним рядом цилиндрических клеток. Жгутиковые стадии отсутствуют.
В центре клеток находится крупное ядро. Оно окружено цитоплазмой, расходящейся в виде тяжей от центра клетки к периферии. Здесь они соединяются с постенным слоем цитоплазмы. Тяжи пронизывают крупную вакуоль. В клетках находятся лентовидные, закрученные в виде спирали хроматофоры. Они располагаются постенно с внутренней стороны оболочки. У разных видов спирогиры количество хроматофоров колеблется от 1 до 16. В хроматофорах в большом количестве располагаются крупные бесцветные пиреноиды. Снаружи водоросль окружена слизистым чехлом.
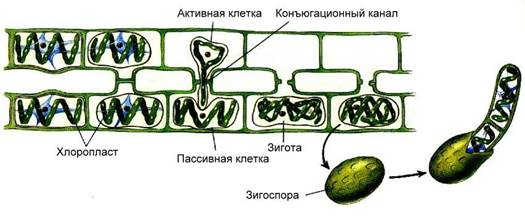
|
| Рис.. Лестничная конъюгация спирогиры |
Рост водоросли в длину осуществляется путем поперечного деления клеток. Размножается спирогира бесполым и половым способом. Бесполое размножение осуществляется частями нитей при их случайном разрыве.
Половой процесс осуществляется путем конъюгации (рис. 64). Конъюгация может быть лестничной и боковой. При лестничной конъюгации две нити располагаются параллельно друг другу. У рядом расположенных клеток образуют куполообразные выросты, растущие навстречу друг другу.
В месте соприкосновения перегородки, разделяющие клетки, растворяются, и образуется канал, связывающий обе клетки. Содержимое одной клетки (мужской) округляется и перетекает по трубке в другую (женскую), и их содержимое (в первую очередь ядра) сливается. При боковой конъюгации оплодотворение происходит в пределах одной нити. При этом наблюдается слияние протопластов двух рядом расположенных клеток.
Зигота, образовавшаяся в результате оплодотворения, окружается толстой клеточной стенкой и впадает в период покоя. Весной зигота редукционно делится и образует четыре гаплоидных ядра. Три ядра дегенерируют, а четвертое делится митотически и дает начало новой гаплоидной нити. Таким образом, спирогира проходит жизненный цикл в гаплоидной фазе, диплоидна у нее только зигота.
Моховидные
Моховидные, общая характеристика. Если у низших растений (водорослей) отсутствовали ткани и органы, то в воздушной среде у псилофитов силурийского периода палеозоя появляются механические, покровные ипроводящие ткани, обеспечивающие возможность жизни в воздушной среде. Появление тканей привело к появлению высших наземных растений, самой примитивной группой среди которых являются моховидные. Считается, что моховидные и сосудистые растения произошли независимо друг от друга от различных групп зеленых водорослей. Родство зеленых водорослей и высших растений подтверждается одинаковым набором фотосинтетических пигментов и накоплением питательных веществ в пластидах, а не в цитоплазме клеток, как у других групп водорослей.
У моховидных, так же как водорослей, корни отсутствуют, их функцию выполняют нитевидные выросты в нижней части стебля – ризоиды. Воду они поглощают слабо, вода захватывается всей поверхностью тела, поэтому они предпочитают места обитания с повышенной влажностью и жизненные формы моховидных – однолетние и многолетние травянистые растения.
Основная особенность, отличающая моховидные растения от высших споровых растений, это преобладание в жизненном цикле гаплоидного гаметофита, на котором развивается диплоидный спорофит. «Стебель» и «листья» мхов – это не настоящие стебель и листья, это образования гаметофита, спорофит (коробочка на ножке) развивается на гаметофите и полностью от него зависит. У всех остальных высших сосудистых растений в жизненном цикле доминирует диплоидный спорофит, гаплоидные гаметофиты все больше редуцируются.
Проводящие ткани – наиболее примитивные среди всех высших растений, настоящая ксилема и флоэмаотсутствуют. Только у наиболее сложно устроенных моховидных появились клетки, напоминающие проводящие ткани ксилемы и флоэмы.
Размножение, как и у водорослей, тесно связано с водой, мужские половые клетки развиваются в мужских гаметангиях гаметофита – антеридиях, яйцеклетки образуются в женских гаметангиях – архегониях. Сперматозоиды по воде добираются до яйцеклеток, происходит оплодотворение и из диплоидной зиготы на гаметофите развивается небольшой спорофит.
Споры образуются путем мейоза и из спор (одноклеточных образований) в благоприятных условиях вновь формируются гаметофиты. Споры морфологически одинаковы, это тоже примитивный признак, все моховидные – равноспоровые растения. Но у многих моховидных споры физиологически неравноценны, из одних развиваются гаметофиты, образующие антеридии, из других – образующие архегонии, то есть у многих моховидных гаметофиты двудомные.
Отдел объединяет около 25 тыс. современных видов и делится на три класса: Антоцеротовые мхи, Печеночники и Листостебельные мхи.
Класс Листостебельные мхи. Кукушкин лен. Кукушкин лен – один из наиболее широко распространенных представителей подкласса Зеленые мхи (рис. 66). Произрастает на влажных местах, в болотах, заболоченных лесах. Это многолетнее растение, достигающее высоты 15-40 см. Произрастает группами, образуя крупные подушковидные дернины."Стебель" мха прямостоячий, неветвящийся. В центре располагаются более вытянутые клетки, соответствующие ксилеме и флоэме. "Стебель" густо покрыт узкими линейно-ланцетными "листьями". Они состоят из нескольких слоев клеток. У основания стебля развиваются многоклеточныенитевидные аналоги корней – ризоиды.
Кукушкин лен относится к двудомным растениям (рис..). На мужском гаметофите, на верхушке, между красноватыми "листочками", образующими розетку, располагаются мужские половые органы – антеридии, в которых образуются двужгутиковые сперматозоиды. Антеридии имеют вид продолговатых или округлых мешочков на ножке. На женском гаметофите образуются женские гаметангии (половые органы) – колбовидные архегонии. В брюшке архегония развивается яйцеклетка. Как и антеридии, архегонии располагаются на верхушке растения. При созревании архегония шейковые и брюшные клетки ослизняются, и на их месте формируется узкий канал, по которому сперматозоиды могут проникнуть к яйцеклетке. Оплодотворение происходит в дождливую погоду, так как для передвижения сперматозоидов необходима водная среда.

|
| Рис. 67. Цикл развития мха кукушкин лен: 1 – гаметофит; 2 – спорофит; 3 – коробочка; 4 – колпачок; 5 – споры; 6 – протонема; 7 – мужской гаметофит; 8 – антеридии; 9 – женский гаметофит; 10 – движение сперматозоидов к архегониям; 11 – архегонии. |
Сперматозоиды обладают положительным хемотаксисом к содержимому слизи архегония, передвигаясь по воде, проникают внутрь архегония, в котором происходит слияние одного из них с яйцеклеткой.
Через несколько месяцев из зиготы прорастает спорофит. Спорофит кукушкина льна состоит из гаустории, ножки и коробочки. Гаустория (присоска) служит для внедрения в тело гаметофита. На ранней стадии спорофит зеленый и способен к фотосинтезу, позднее желтеет, затем становится оранжевым и, наконец, бурым и полностью переходит к питанию за счет гаметофита. На верхнем конце коробочки до созревания находится колпачок, калиптра. Он развивается из стенки брюшка архегония и остается гаплоидным. В коробочках путем мейотического деления происходит образование спор (спорическая редукция). Все споры морфологически одинаковы, но физиологически отличаются.
После созревания колпачок и крышечка отпадают, и споры легко рассеиваются с помощью ветра. При благоприятных условиях спора прорастает в тонкую ветвящуюся зеленую нить – протонему, или предросток. С нее начинается развитие нового гаметофита. На протонеме образуются почки, из которых развиваются гаметофиты – взрослые растения мха, какого-либо одного пола, имеющие гаплоидный набор хромосом. Таким образом, у кукушкина льна, как и всех остальных моховидных, да и для всех остальных наземных растений, характерно чередование гаплоидного (n) и диплоидного (2n) поколений (гаметофита и спорофита). Спорофит, как правило, паразитирует на гаметофите (исключение составляют Антоцеротовые).
Торфяной мох сфагнум. К сфагновым мхам относится свыше 300 видов единственного рода сфагнум, распространенных преимущественно на севере Евразии и Америки. Здесь они занимают обширные площади, являясь основными образователями торфяных болот.
Мох сфагнум – небольшие растение (до 15—20 см), белесоватого цвета, боковые побеги которого густо покрыты узкими длинными листьями (рис. 68). Растет обычно плотными дернинами. Стебель взрослого растения ризоидов не имеет. Он ежегодно нарастает верхушкой, в то время как его нижняя часть постоянно отмирает. Спрессованные слои отмершего сфагнума образуют залежи торфа.
Листья сфагнума яйцевидной формы, без средней жилки. Они образованы одним слоем клеток двух типов: узкими длинными живыми, содержащих хлоропласты – ассимилирующие, образующие как бы сетку и широкими мертвыми гиалиновыми водоносными клетками со спиральными утолщениями, располагающимися между живыми.
Мертвые клетки имеют отверстия, поры и способны накапливать и удерживать большое количество воды (в 25—37 раз больше своего веса).

|
| Рис.. Сфагновый мох: 1 – вертикальный побег; 2 – горизонтальный побег; 3 – коробочка со спорами; 4 – ассимилирующая (хлорофиллоносная) клетка; 5 – гиалиновая (водоносная) клетка; б – пора; 7 – ложная ножка; 8 – гаустория; 9 – молодая протонема. |
Сфагнум – однодомное растение, антеридии и архегонии формируются на боковых веточках в верхней части стебля. Оплодотворение яйцеклеток двужгутиковыми сперматозоидами происходит при наличии воды.
Из зиготы развивается спорофит, представленный круглой коробочкой. Гаустория спорофита врастает в подставку из тканей гаметофита – ложную ножку.
К моменту созревания спор (в результате мейоза) подставки удлиняются и коробочки поднимаются над облиственной частью стебля.
Во влажную погоду через устьица проникает воздух, при подсыхании коробочки устьица закрываются, давление в коробочке повышается и с отчетливым хлопком крышечка срывается, и облачко спор поднимается над коробочкой. Попав в благоприятные условия, споры прорастают в однослойную пластинчатую протонему, на которой возникают почки, дающие начало новым побегам мха.
Сфагнум в четыре раза гигроскопичнее ваты и содержит вещество – сфагнол, обладающее бактерицидным действием. Кроме того, сфагнум не только заболачивает, но и подкисляет почву до рН ниже 4. В кислой бактерицидной среде бактерии гниения погибают, а остатки растений оседают на дно и спрессовываются, превращаясь в торф.
Значение мхов. Моховидные в природе часто поселяются на таких субстратах и в таких местообитаниях, которые недоступны для других растений. В этом случае они выступают в роли пионерной растительности, играя большую роль в почвообразовательных процессах. Моховидные играют существенную роль в регуляции водного баланса суши. Они регулируют испарение влаги из почвы.
На лугах мхи препятствуют семенному возобновлению трав, в лесах – прорастанию семян деревьев. Аккумулируя воду, мхи вызывают заболачивание почв. Сфагновые и зеленые мхи являются основными торфообразователями. Наличие мохового покрова является одним из основных стабилизирующих факторов в условиях вечной мерзлоты.
Хозяйственное значение. Животные мхи не поедают. Торф применяют в качестве топлива, подстилки для домашних животных, удобрения. Путем сухой перегонки торфа получают метиловый спирт, сахарин, воск, парафин, краски, и т.д. Из торфа изготавливают бумагу и картон. В строительстве торф используется как теплоизолирующий и звукоизолирующий материал. Сфагнум имеет и медицинское значение – используется как прекрасный перевязочный материал.
Папоротникообразные
Самые древние сосудистые растения на Земле – риниофиты. Они появились в силурийском периоде палеозойской эры, около 440 млн. лет назад и росли в прибрежной зоне. Настоящих корней они еще не имели, в почве находился горизонтальный побег, от которого поднимались вверх вертикальные, дихотомически ветвящиеся оси, многие из которых заканчивались спорангиями. Все риниофиты были равноспоровыми растениями. Листья еще отсутствовали, роль корней выполняли ризоиды. Но это уже были сосудистые растения, у них уже сформировалась ксилема, проводящая воду вверх по стеблю и флоэма, проводящая органические вещества, окружала центральный тяж ксилемы. Центральный проводящий пучок был окружен механической тканью и клетками коры, снаружи уже была покровная ткань – эпидерма, имеющая устьица. Механическая, проводящая и покровная ткани позволили растениям приспособиться к жизни в воздушной среде и начать освоение суши.
 Рис..Риниевые а) риния, внешний вид; б) поперечный срез стебля; в) продольный разрез спорангия; 1 – ризоиды; 2 – спорангии; 3 –эпидерма; 4 – устьице; 5 – кора; 6 – флоэма; 7 – ксилема. Рис..Риниевые а) риния, внешний вид; б) поперечный срез стебля; в) продольный разрез спорангия; 1 – ризоиды; 2 – спорангии; 3 –эпидерма; 4 – устьице; 5 – кора; 6 – флоэма; 7 – ксилема.
|
Дальнейшее освоение суши сопровождалось появлением корней и листьев. От одной из групп риниофитов (зостерофиллофитов) произошли плауновидные, причем листья у них образовывались как уплощенные боковые стебли с единственной жилкой (проводящим пучком), такие листья
|
|
|
