 |


Структура дезоксирибонуклеиновой кислоты (ДНК)
|
|
|
|
ЛАБОРАТОРНАЯ РАБОТА №2
Тема: СТРОЕНИЕ НУКЛЕИНОВЫХ КИСЛОТ
Цель работы: изучить химическую, структурную и пространственную организацию нуклеиновых кислот
Общая информация. В каждом живом организме присутствуют 2 типа нуклеиновых кислот: рибонуклеиновая кислота (РНК) и дезоксирибонуклеиновая кислота (ДНК). Измерения размера генома приводятся в дальтонах, парах нуклеотидов (п. н.) или пикограммах (пг). Соотношение между этими единицами измерения следующие:
1 пг = 10–9 мг = 0,6х1012 дальтон = 0,9х109 п. н.
В гаплоидном геноме человека содержится около 3,2 млрд. п. н., что равно 3,5 пг ДНК. Таким образом, в ядре одной клетки человека содержится около 7 пг ДНК. Если учесть, что средний вес клетки человека равен примерно 1000 пг, то легко рассчитать, что ДНК составляет менее 1% от веса клетки. Молекулярная масса самой «маленькой» из известных нуклеиновых кислот - транспортной РНК (тРНК) составляет примерно 25 кД. ДНК - наиболее крупные полимерные молекулы; их молекулярная масса варьирует от 1 000 до 1 000 000 кД. ДНК и РНК состоят из мономерных единиц - нуклеотидов, поэтому нуклеиновые кислоты называют полинуклеотидами.
Строение нуклеотидов
Каждый нуклеотид содержит 3 химически различных компонента: гетероциклическое азотистое основание, моносахарид (пентозу) и остаток фосфорной кислоты. В зависимости от числа имеющихся в молекуле остатков фосфорной кислоты различают нуклеозидмонофосфаты (НМФ), нуклеозиддифосфаты (НДФ), нуклеозидтрифосфаты (НТФ) (рис. 1).
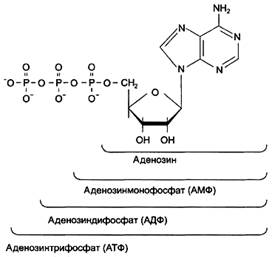
Рисунок 1. Нуклеозидмоно-, ди- и трифосфаты аденозина. Нуклеотиды - фосфорные эфиры нуклеозидов. Остаток фосфорной кислоты присоединён к 5'-углеродному атому пентозы (5'-фосфоэфирная связь).
|
|
|
В состав нуклеиновых кислот входят азотистые основания двух типов: пуриновые - аденин (А),гуанин (G) и пиримидиновые - цитозин (С), тимин (Т) и урацил (U). Нумерация атомов в основаниях записывается внутри цикла (рис. 2). Номенклатура нуклеотидов приведена в табл. 1.
Таблица 1. Номенклатура нуклеотидов
| Азотистое основание | Нуклеозид | Нуклеотид | Трёхбуквенное обозначение | Однобуквенный код |
| Аденин | Аденозин | Аденозинмонофосфат | АМФ | А |
| Гуанин | Гуанозин | Гуанозинмонофосфат | ГМФ | G |
| Цитозин | Цитидин | Цитидинмонофосфат | ЦМФ | С |
| Урацил | Уридин | Уридинмонофосфат | УМФ | U |
| Тимин | Тимидин | Тимидинмонофосфат | ТМФ | Т |
Пентозы в нуклеотидах представлены либо рибозой (в составе РНК), либо дезоксирибозой (в составе ДНК). Чтобы отличить номера атомов в пентозах от нумерации атомов в основаниях, запись производят с внешней стороны цикла и к цифре добавляют штрих (') - 1', 2', 3', 4' и 5' (рис. 3).
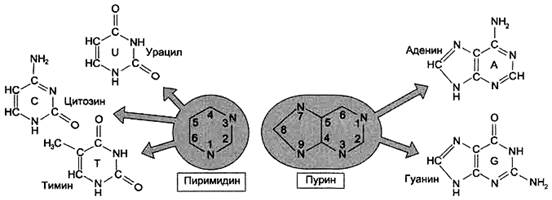
Рисунок 2. Пуриновые и пиримидиновые основания.
Пентозу соединяет с основанием N-гликозидная связь, образованная С1-атомом пентозы (рибозы или дезоксирибозы) и N1 -атомом пиримидина или N9-aтомом пурина (рис. 4).
Нуклеотиды, в которых пентоза представлена рибозой, называют рибонуклеотидами, а нуклеиновые кислоты, построенные из рибонуклеотидов, - рибонуклеиновыми кислотами, или РНК. Нуклеиновые кислоты, в мономеры которых входит дезоксирибоза, называют дезоксирибонуклеиновыми кислотами, или ДНК. Нуклеиновые кислоты по своему строению относят к классу линейных полимеров.
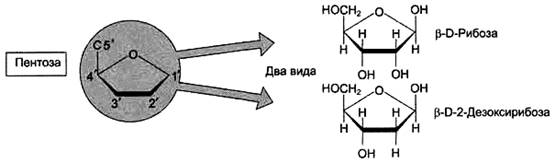
Рисунок 3. Пентозы. Присутствуют 2 вида - β-D-рибоза в составе нуклеотидов РНК и β-D-2-дезоксирибоза в составе нуклеотидов ДНК.
Остов нуклеиновой кислоты имеет одинаковое строение по всей длине молекулы и состоит из чередующихся групп - пентоза-фосфат-пентоза- (рис. 5). Вариабельными группами в полинуклеотидных цепях служат азотистые основания - пурины и пиримидины. В молекулы РНК входят аденин (А), урацил (U), гуанин (G) и цитозин (С), в ДНК - аденин (А), тимин (Т), гуанин (G) и цитозин (С). Уникальность структуры и функциональная индивидуальность молекул ДНК и РНК определяются их первичной структурой - последовательностью азотистых оснований в полинуклеотидной цепи.
|
|
|
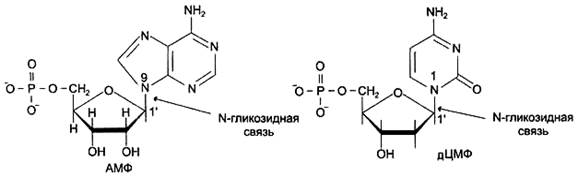
Рисунок 4. Пуриновый и пиримидиновый нуклеотиды.
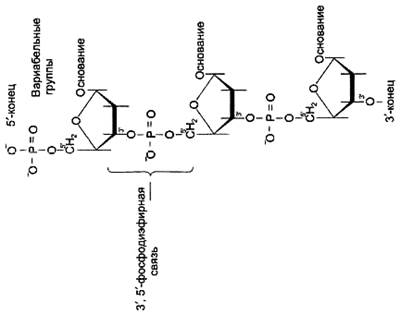
Рисунок 5. Фрагмент цепи ДНК.
Структура дезоксирибонуклеиновой кислоты (ДНК)
Первичная структура ДНК - порядок чередования дезоксирибонуклеозидмонофосфатов (дНМФ) в полинукпеотидной цепи.
Каждая фосфатная группа в полинукпеотидной цепи, за исключением фосфорного остатка на 5'-конце молекулы, участвует в образовании двух эфирных связей с участием 3'- и 5'-углеродных атомов двух соседних дезоксирибоз, поэтому связь между мономерами обозначают 3', 5'-фосфодиэфирной.
Концевые нуклеотиды ДНК различают по структуре: на 5'-конце находится фосфатная группа, а на 3'-конце цепи - свободная ОН-группа. Эти концы называют 5'- и 3'-концами. Линейная последовательность дезоксирибонуклеотидов в полимерной цепи ДНК обычно сокращённо записывают с помощью однобуквенного кода, например -A-G-C-T-T-A-C-A- от 5'- к 3'-концу.
В каждом мономере нуклеиновой кислоты присутствует остаток фосфорной кислоты. При рН 7 фосфатная группа полностью ионизирована, поэтому in vivo нуклеиновые кислоты существуют в виде полианионов (имеют множественный отрицательный заряд). Остатки пентоз тоже проявляют гидрофильные свойства. Азотистые основания почти нерастворимы в воде, но некоторые атомы пуринового и пиримидинового циклов способны образовывать водородные связи.
Вторичная структура ДНК. В 1953 г. Дж. Уотсоном и Ф. Криком была предложена модель пространственной структуры ДНК. Согласно этой модели, молекула ДНК имеет форму спирали, образованную двумя полинуклеотидными цепями, закрученными относительно друг друга и вокруг общей оси. Двойная спираль правозакрученная, полинуклеотидньхе цепи в ней антипараллельны (рис. 6), т.е. если одна из них ориентирована в направлении 3'→5', то вторая - в направлении 5'→3'. Поэтому на каждом из концов молекулы ДНК расположены 5'-конец одной цепи и 3'-конец другой цепи.
|
|
|
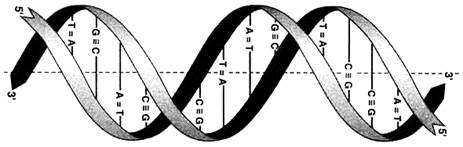
Рисунок 6. Двойная спираль ДНК. Молекулы ДНК состоят из двух антипараллельных цепей с комплементарной последовательностью нуклеотидов. Цепи закручены относительно друг друга в правозакрученную спираль так, что на один виток приходится примерно 10 пар нуклеотидов.
Все основания цепей ДНК расположены внутри двойной спирали, а пентозофосфатный остов - снаружи. Полинуклеотидные цепи удерживаются относительно друг друга за счёт водородных связей между комплементарными пуриновыми и пиримидиновыми азотистыми основаниями А и Т (две связи) и между G и С (три связи) (рис. 7, 8).
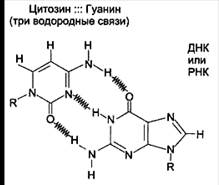
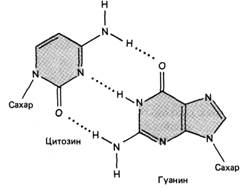


Рисунок 7. Пурин-пиримидиновые пары оснований в ДНК.
При таком сочетании каждая пара содержит по три кольца, поэтому общий размер этих пар оснований одинаков по всей длине молекулы. Водородные связи при других сочетаниях оснований в паре возможны, но они значительно слабее. Последовательность нуклеотидов одной цепи полностью комплементарна последовательности нуклеотидов второй цепи. Поэтому, согласно правилу Чаргаффа (Эрвин Чаргафф в 1951 г. установил закономерности в соотношении пуриновых и пиримидиновых оснований в молекуле ДНК), число пуриновых оснований (А + G) равно числу пиримидиновых оснований (Т + С).
5΄ фосфат 3΄ гидроксильная группа
3΄ гидроксильная группа 5΄ фосфат
Рисунок 8. Из-за противоположной последовательности атомов в цепях обе цепи инвертированы относительно друг друга, т.е. направления от 3'-конца к 5'-концу в двух цепях, из которых состоит молекула ДНК, противоположны (цепи «антипараллельны» друг другу).
Комплементарые основания уложены в стопку в сердцевине спирали. Между основаниями двухцепочечной молекулы в стопке возникают гидрофобные взаимодействия, стабилизирующие двойную спираль.
Такая структура исключает контакт азотистых остатков с водой, но стопка оснований не может быть абсолютно вертикальной. Пары оснований слегка смещены относительно друг друга. В образованной структуре различают две бороздки - большую, шириной 2,2 нм, и малую, шириной 1,2 нм. Азотистые основания в области большой и малой бороздок взаимодействуют со специфическими белками, участвующими в организации структуры хроматина.
|
|
|
Водородные связи между основаниями двух нитей могут разрываться. Разрыв водородных связей может происходить под воздействием различных факторов, что приводит к денатурации молекулы. Например, нагревание ДНК до 950 С вызывает разрушение водородных связей. Это явление называется плавлением ДНК.
Цепочки двойной спирали могут расходиться как замок-молния и под действием ферментов (хеликазы). Причем, на разрыв ГЦ требуется больше энергии, чем на разрыв пары АТ. Процент ГЦ пар и длина молекулы ДНК определяют количество энергии, необходимой для диссоциации цепей: длинные молекулы ДНК с большим содержанием ГЦ более тугоплавки. Поэтому, части молекул ДНК, которые из-за их функций должны быть легко разделяемы, например, последовательности в бактериальных промоторах, обычно содержат большое количество А и Т.
Третичная структура ДНК (суперспирализация ДНК) определяется трехмерной пространственной организацией молекул и зависит от различных факторов. Компактизация и суперспирализация ДНК осуществляются с помощью разнообразных белков, взаимодействующих с определёнными последовательностями в структуре ДНК. Все связывающиеся с ДНК эукариотов белки можно разделить на 2 группы: гистоновые и негистоновые белки. Комплекс белков с ядерной ДНК клеток называют хроматином.
|
|
|
