 |


Практическая работа №6. Межклеточные мембранные контакты. Производные плазмолеммы. Межклеточные контакты
|
|
|
|
Практическая работа №6
Межклеточные мембранные контакты.
Производные плазмолеммы
Цель: Изучить межклеточные мембранные контакты и производные плазмолеммы.
¨ Задание:
1. Прочитайте статью «Межклеточные контакты» и заполните таблицу 7 «Типы контактов».
Таблица 7
Типы контактов
| № | Тип контакта | Характеристика | Рисунок |
| 1. | Плотный замыкающий контакт | ||
| 2. | Простой контакт | ||
| 3. | Зубчатый контакт («замок») | ||
| 4. | Десмосомы; полудесмосомы | ||
| 6. | Плотный замыкающий контакт | ||
| 7. | Щелевые контакты (нексусы) | ||
| 8. | Синаптический контакт | ||
| 9. | Плазмодесмы |
2. Изучите и зарисуйте строение производных плазмолеммы:
а) Микроворсинки.

| 
|
|
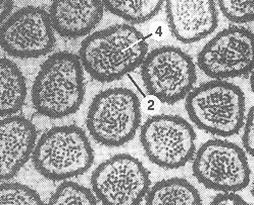
Рис. 7. Микрофотография микроворсинок. 1-микроворсинки: цилиндрические пальцеобразные выросты цитоплазмы; 2 – плазмолемма; 3 – просвет тонкой кишки; 4 – микрофиламенты, расположенные вдоль оси микроворсинок. | |
б) Щеточная каемка.
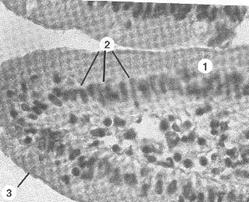
| Рис. 8. Микрофотографии щеточной каемки клеток тонкой кишки. 1 – ворсинка, образованная слизистой оболочкой тонкой кишки; 2 – клетки (цилиндрической формы), покрывающие ворсинки; 3 – щеточная оксифильная каемка на апикальной поверхности клеток, образованная микроворсинками. |
Межклеточные контакты
У многоклеточных организмов за счет межклеточных взаимодействий образуются сложные клеточные ансамбли, поддержание которых может осуществляться разными путями. В зародышевых, эмбриональных тканях, особенно на ранних стадиях развития, клетки остаются в связи друг с другом за счет способности их поверхностей слипаться. Это свойство адгезии (соединения, сцепления) клеток может определяться свойствами их поверхности, которые специфически взаимодействуют друг с другом. Механизм этих связей достаточно хорошо изучен, он обеспечивается взаимодействием между гликопротеидами плазматических мембран.
|
|
|
Кроме сравнительно простых адгезивных (но специфических) связей существует целый ряд специальных межклеточных структур, контактов или соединений, которые выполняют определенные функции.
Запирающее или плотное соединение характерно для однослойных эпителиев (Рис. 9). Это зона, где внешние слои двух плазматических мембран максимально сближены. Часто видна трехслойность мембраны в этом контакте: два внешних осмофильных слоя обеих мембран как бы сливаются в один общий слой толщиной 2-3 нм.
Слияние мембран происходит не по всей площади плотного контакта, а представляет собой ряд точечных сближений мембран. Такие структуры при специальных окрасках можно видеть и в световом микроскопе. Они получили у морфологов название замыкающих пластинок. Роль замыкающего плотного контакта заключается не только в механическом соединении клеток друг с другом. Эта область контакта плохо проницаема для макромолекул и ионов, и тем самым она запирает, перегораживает межклеточные полости, изолируя их (и вместе с ними собственно внутреннюю среду организма) от внешней среды (в данном случае - просвет кишечника).
Замыкающий, или плотный, контакт встречается между всеми типами однослойного эпителия (эндотелий, мезотелий, эпендима).
Простой контакт, встречающийся среди большинства прилегающих друг к другу клеток различного происхождения (Рис. 10). Большая часть поверхности контактирующих клеток эпителия также связана с помощью простого контакта, где плазматические мембраны, соприкасающихся клеток разделены пространством 15-20 нм. Это пространство представляет собой надмембранные компоненты клеточных поверхностей. Ширина щели между мембранами клеток может быть и больше 20 нм, образуя расширения, полости, но не меньше 10 нм.
|
|
|
Со стороны цитоплазмы к этой зоне плазматической мембраны не примыкают никакие специальные дополнительные структуры.
Зубчатый контакт («замок») представляет собой выпячивание поверхности плазматической мембраны одной клетки в инвагинат (впячивание) другой (Рис. 11).
На срезе такой тип соединения напоминает плотничий шов. Межмембранное пространство и цитоплазма в зоне «замков» имеют те же характеристики, что и в зонах простого контакта. Такой тип межклеточных соединений характерен для многих эпителиев, где он соединяет клетки в единый пласт, способствуя, их механическому скреплению друг с другом.
Роль механического плотного скрепления клеток друг с другом играет ряд специальных структурированных межклеточных соединений.
Десмосомы, структуры в виде бляшек или кнопок также соединяют клетки друг с другом (Рис. 12). В межклеточном пространстве здесь также виден плотный слой, представленный взаимодействующими интегральными мембранными кадгеринами - десмоглеинами, которые сцепляют клетки друг с другом.
С цитоплазматической стороны к плазмолемме прилежит слой белка-десмоплакина, с которым связаны промежуточные филаменты цитоскелета. Десмосомы встречаются чаще всего в эпителиях, в этом случае промежуточные филаменты содержат кератины. В сердечной мышце клетки, кардиомиоциты, содержат десминовые фибриллы в составе десмосом. В эндотелии сосудов в состав десмосом входят виментиновые промежуточные филаменты.
Полудесмосомы, в принципе, сходны по строению с десмосомой, но представляют собой соединение клеток с межклеточными структурами. Так в эпителиях линкерные гликопротеиды (интегрины) десмосомы взаимодействуют с белками т. н. базальной мембраны, куда входят коллаген, ламинин, протеогликаны и др.
Функциональная роль десмосом и полудесмосом сугубо механическая - они сцепляют клетки друг с другом и с подлежащим внеклеточным матриксом прочно, что позволяет эпителиальным пластам выдерживать большие механические нагрузки.
|
|
|
Подобно этому десмосомы прочно связывают друг с другом клетки сердечной мышцы, что позволяет им выполнять огромную механическую нагрузку, оставаясь связанными в единую сокращающуюся структуру.
В отличие от плотного контакта все типы сцепляющих контактов проницаемы для водных растворов и не играют никакой роли в ограничении диффузии.
Щелевые контакты (нексусы) считаются коммуникационными соединениями клеток; это структуры, которые участвуют в прямой передаче химических веществ из клетки в клетку, что может играть большую физиологическую роль не только при функционировании специализированных клеток, но и обеспечивать межклеточные взаимодействия при развитии организма, при дифференцировке его клеток (Рис. 13).
Характерным для этого типа контактов является сближение плазматических мембран двух соседних клеток на расстояние 2-3 нм. Именно это обстоятельство долгое время не позволяло на ультратонких срезах отличить данный вид контакта от плотного разделительного (замыкающего) контакта. При использовании гидроокиси лантана было замечено, что некоторые плотные контакты пропускают контрастер. В этом случае лантан заполнял тонкую щель шириной около 3 нм между сближенными плазматическими мембранами соседних клеток. Это и послужило появлению термина - щелевой контакт. Дальнейший прогресс в расшифровке его строения был достигнут при использовании метода замораживания-скалывания. Оказалось, что на сколах мембран зоны щелевых контактов (размеров от 0, 5 до 5 мкм) усеяны гексагонально расположенными с периодом 8-10 нм частицами 7-8 нм в диаметре, имеющими в центре канал около 2 нм шириной. Эти частицы получили название коннексонов.
В зонах щелевого контакта может быть от 10-20 до нескольких тысяч коннексонов в зависимости от функциональных особенностей клеток. Коннексоны были выделены препаративно, они состоят из шести субъединиц коннектина - белка с молекулярным весом около 30 тыс. Объединяясь друг с другом, коннектины образуют цилиндрический агрегат – коннексон, в центре которого располагается канал.
|
|
|
Отдельные коннексоны встроены в плазматическую мембрану так, что прободают ее насквозь. Одному коннексону на плазматической мембране клетки точно противостоит коннексон на плазматической мембране соседней клетки так, что каналы двух коннексонов образуют единое целое. Коннексоны играют роль прямых межклеточных каналов, по которым ионы и низкомолекулярные вещества могут диффундировать из клетки в клетку. Было обнаружено, что коннексоны могут закрываться, изменяя диаметр внутреннего канала, и тем участвовать в регуляции транспорта молекул между клетками.
Функциональное значение щелевых контактов было понято при изучении гигантских клеток слюнных желез двукрылых. В такие клетки благодаря их величине легко можно вводить микроэлектроды, для того чтобы изучать электропроводимость их мембран. Если ввести электроды в две соседние клетки, то их плазматические мембраны проявляют низкое электрическое сопротивление, между клетками идет ток. Такая способность щелевых контактов служить местом транспорта низкомолекулярных соединений используется в тех клеточных системах, где нужна быстрая передача электрического импульса (волны возбуждения) от клетки к клетке без участия нервного медиатора. Так, все мышечные клетки миокарда сердца связаны с помощью щелевых контактов (кроме того, клетки там связаны и адгезивными контактами). Это создает условие для синхронного сокращения огромного количества клеток.
При росте культуры эмбриональных сердечных мышечных клеток (кардиомиоциты) некоторые клетки в пласте начинают независимо друг от друга спонтанно сокращаться с разной частотой, и лишь только после образования между ними щелевых контактов они начинают биться синхронно как единый сокращающийся пласт клеток. Таким же способом обеспечивается совместное сокращение гладкомышечных клеток в стенке матки.
Синаптический контакт (синапсы). Этот тип контактов характерен для нервной ткани и встречается как между двумя нейронами, так и между нейроном и каким-либо иным элементом – рецептором или эффектором (например, нервно-мышечное окончание) (Рис. 14).

| 
|
| Рис. 9. Плотный контакт | Рис. 10. Простой контакт |

| 
|
| Рис. 11. Зубчатый контакт | Рис. 12. Десмосомы |

| 
|
| Рис. 13. Нексусы | Рис. 14. Синаптический контакт |
Синапсы – участки контактов двух клеток, специализированных для односторонней передачи возбуждения или торможения от одного элемента к другому. В принципе подобного рода функциональная нагрузка, передача импульса может осуществляться и другими типами контактов (например, щелевым контактом в сердечной мышце), однако в синаптической связи достигается высокая эффективность в реализации нервного импульса.
|
|
|
Синапсы образуются на отростках нервных клеток – это терминальные участки дендритов и аксонов. Межнейронные синапсы обычно имеют грушевидных расширений, бляшек на конце отростка нервной клетки. Такое терминальное расширение отростка одной из нервных клеток может контактировать и образовывать синаптическую связь как с телом другой нервной клетки, так и с ее отростками. Периферические отростки нервных клеток (аксоны) образуют специфические контакты с клетками-эффекторами или клетками-рецепторами. Следовательно, синапс - это структура, образующаяся между участками двух клеток (так же как и десмосома). Мембраны этих клеток разделены межклеточным пространством - синаптической щелью шириной около 20-30 нм. Часто в просвете этой щели виден тонковолокнистый, перпендикулярно расположенный по отношению к мембранам материал. Мембрана в области синаптического контакта одной клетки называется пресинаптической, другой, воспринимающей импульс, - постсинаптической. В электронном микроскопе обе мембраны выглядят плотными, толстыми. Около пресинаптической мембраны выявляется огромное количество мелких вакуолей, синаптических пузырьков, заполненных медиаторами. Синаптические пузырьки в момент прохождения нервного импульса выбрасывают свое содержимое в синаптическую щель. Постсинаптическая мембрана часто выглядит толще обычных мембран из-за скопления около нее со стороны цитоплазмы множества тонких фибрилл.
Плазмодесмы. Этот тип межклеточных связей встречается у растений. Плазмодесмы представляют собой тонкие трубчатые цитоплазматические каналы, соединяющие две соседние клетки (Рис. 15). Диаметр этих каналов обычно составляет 20-40 нм. Ограничивающая эти каналы мембрана непосредственно переходит в плазматические мембраны соседствующих клеток.
Плазмодесмы проходят сквозь клеточную стенку, разделяющую клетки. Таким образом, у некоторых растительных клеток плазмодесмы соединяют гиалоплазму соседних клеток, поэтому формально здесь нет полного разграничения, отделения тела одной клетки от другой, это скорее представляет собой синцитий: объединение многих клеточных территорий с помощью цитоплазматических мостиков.
Внутрь плазмодесм могут проникать мембранные трубчатые элементы, соединяющие цистерны эндоплазматического ретикулума соседних клеток. Образуются плазмодесмы во время деления клетки, когда строится первичная клеточная оболочка. У только что разделившихся клеток число плазмодесм может быть очень велико (до 1000 на клетку), при старении клеток их число падает за счет разрывов при увеличении толщины клеточной стенки.
Функциональная роль плазмодесм очень велика: с их помощью обеспечивается межклеточная циркуляция растворов, содержащих питательные вещества, ионы и другие соединения.
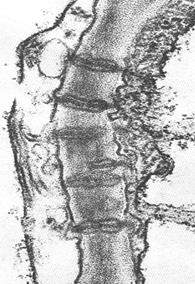
| По плазмодесмам могут перемещаться липидные капли. Через плазмодесмы происходит заражение клеток растительными вирусами. Однако эксперименты показывают, что свободный транспорт через плазмодесмы ограничивается частицами с массой не более 800 дальтон. Рис. 15. Плазмодесмы |
|
|
|
