 |


Гисто- и органогенез. Процесс нейруляции. Осевые органы и их формирование. Дифференцировка мезодермы. Провизорные органы зародышей позвоночных.
|
|
|
|
Гомология зародешевых листков. Производные зародешевых листков.
По окончании периода дробления у многоклеточных животных начинается период образования зародешевых листков – гаструляция. Гаструляция связана с перемещением эмбрионального материала. Гаструла (гр. гастер – желудок): зародыш двухслойный, у него появляется кишечная полость, первичное ротовое отверстие, два слоя клеток – эктодерма и энтодерма. Затем следует стадия поздней гаструлы (у всех животных, кроме губок и кишечнополостных). На этой стадии появляется третий слой клеток – мезодерма, которая закладывается между экто- и энтодермой. Вначале она имеет вид двух карманов, полости которых представляют собой вторичную полость тела.
Зародышевые листки - (зародышевые пласты) - слои тела зародыша многоклеточных животных и человека, образующиеся в процессе гаструляции. У большинства организмов три зародышевых листка: наружный - эктодерма, внутренний - энтодерма и средний - мезодерма. Каждый зародышевый листок дает начало определенной группе тканей и органов.
В дальнейшем идет дифференцировка клеток: из эктодермы образуются наружный эпителий, кожные железы, эмаль зубов, волосы, ногти. Из энтодермы – эпителий средней кишечники, печень, поджелудочная железа, щитовидная железа, тимус, эпителий дыхательной системы. Из мезодермы – скелет, мышцы, кровеносная система, каналов выделительной системы, части тканей половых желез. У всех животных и у человека одни и те же зародышевые листки формируют одни и те же органы и ткани. Это является свидетельством того, что зародышевые листки гомологичны и имеют единое происхождение в эволюции. Дальнейшее развитие зародыша идет в строгой зависимости одних органов от других (закон эмбриональной индукции Г.Шпемана).
|
|
|
Гисто- и органогенез. Процесс нейруляции. Осевые органы и их формирование. Дифференцировка мезодермы. Провизорные органы зародышей позвоночных.
После образования мезодермы начинается процесс гисто- и органогенеза. В ходе органогенезов изменяются форма, структура и химический состав клеток, обособляются клеточные группы, представляющие собой зачатки будущих органов. Обязательным условием органогенезов наряду с размножением, миграцией и сортировкой клеток является их избирательная гибель.
Самое начало органогенеза называют нейруляцией. В процессе нейруляции возникает комплекс осевых органов нервная трубка — хорда — кишка. Нейруляция охватывает процессы от появления первых признаков формирования нервной пластинки до замыкания ее в нервную трубку. Параллельно формируются хорда и вторичная кишка, а лежащая по бокам от хорды мезодерма расщепляется в краниокаудальном направлении на сегментированные парные структуры — сомиты.
Вначале неспециализированная спинная эктодерма, отвечая на индукционное воздействие со стороны хордомезодермы, превращается в нервную пластинку, представленную нейроэпителиальными клетками цилиндрической формы.
Нервная пластинка недолго остается уплощенной. Вскоре ее боковые края приподнимаются, образуя нервные валики, которые лежат по обе стороны неглубокой продольной нервной бороздки. Края нервных валиков далее смыкаются, образуя замкнутую нервную трубку с каналом внутри — невроцелем. Раньше всего смыкание нервных валиков происходит на уровне начала спинного мозга, а затем распространяется в головном и хвостовом направлениях. Показано, что в морфогенезе нервной трубки большую роль играют микротрубочки и микрофиламенты нейроэпителиальных клеток. Разрушение этих клеточных структур колхицином и цитохалазином В приводит к тому, что нервная пластинка остается открытой. Несмыкание нервных валиков ведет к врожденным порокам развития нервной трубки.
|
|
|
После смыкания нервных валиков клетки, первоначально располагавшиеся между нервной пластинкой и будущей кожной эктодермой, образуют нервный гребень. Клетки нервного гребня отличаются способностью к обширным, но строго регулируемым миграциям по всему и образуют два главных потока. Клетки одного из них—поверхностного—включаются в эпидермис или дерму кожи, где дифференцируются в пигментные клетки. Другой поток мигрирует в брюшном направлении, образует чувствительные спинномозговые ганглии, симпатические нервные узлы, мозговое вещество надпочечников, парасимпатические ганглии. Клетки из черепного отдела нервного гребня дают начало как нервным клеткам, так и ряду других структур, таких, как жаберные хрящи, некоторые кроющие кости черепа.
Мезодерма, занимающая место по бокам от хорды и распространяющаяся далее между кожной эктодермой и энтодермой вторичной кишки, подразделяется на дорсальную и вентральную области. Дорсальная часть сегментирована и представлена парными сомитами. Закладка сомитов идет от головного к хвостовому концу. Вентральная часть мезодермы, имеющая вид тонкого слоя клеток, называется боковой пластинкой. Сомиты соединены с боковой пластинкой промежуточной мезодермой в виде сегментированных ножек сомитов.
Все области мезодермы постепенно дифференцируются. В начале формирования сомиты имеют конфигурацию, характерную для эпителия с полостью внутри. Под индукционным воздействием, исходящим от хорды и нервной трубки, вентромедиальные части сомитов — склеротомы —превращаются во вторичную мезенхиму, выселяются из сомита и окружают хорду и вентральную часть нервной трубки. В конце концов из них образуются позвонки, ребра и лопатки.
Дорсолатеральная часть сомитов с внутренней стороны образует миотомы, из которых разовьются поперечно-полосатые скелетные мышцы тела и конечностей. Наружная дорсолатеральная часть сомитов образует дерматомы, которые дают начало внутреннему слою кожи — дерме. Из области ножек сомитов с зачатками нефротом и гонотом образуются органы выделения и половые железы.
|
|
|
Правая и левая несегментированные боковые пластинки расщепляются на два листка, ограничивающих вторичную полость тела — целом. Внутренний листок, прилежащий к энтодерме, называют висцеральным. Он окружает кишку со всех сторон и образует брыжейку, покрывает легочную паренхиму и мышцу сердца. Наружный листок боковой пластинки прилежит к эктодерме и называется париетальным. В дальнейшем он образует наружные листки брюшины, плевры и перикарда.
Мезодерма помимо скелетных структур, скелетной мускулатуры, дермы кожи, органов выделительной и половой систем образует сердечно-сосудистую систему, лимфатическую систему, плевру, брюшину и перикард. Из мезенхимы, имеющей смешанное происхождение за счет клеток трех зародышевых листков, развиваются все виды соединительной ткани, гладкая мускулатура, кровь и лимфа.
Провизорные (временные) органы – появляются для осуществления связи зародыша со средой: желточный мешок, амнион, аллантоис, хорион, плацента и серозная оболочка. Цель: обеспечение жизненных функций зародыша.
Амнион представляет собой эктодермальный мешок, заключающий зародыша и заполненный амниотической жидкостью. Амниотическая оболочка специализирована для секреции и поглощения амниотической жидкости, омывающей зародыш. Амнион играет первостепенную роль в защите зародыша от высыхания и от механических повреждений, создавая для него наиболее благоприятную и естественную водную среду. Амнион имеет и мезодермальный слой из внезародышевой соматоплевры, который дает начало гладким мышечным волокнам. Сокращения этих мышц вызывают пульсацию амниона, а медленные колебательные движения, сообщаемые при этом зародышу, по-видимому, способствуют тому, что его растущие части не мешают друг другу.
Хорион (сероза) — самая наружная зародышевая оболочка, прилежащая к скорлупе или материнским тканям, возникающая, как и амнион, из эктодермы и соматоплевры. Хорион служит для обмена между зародышем и окружающей средой. У яйцекладущих видов основная его функция — дыхательный газообмен; у млекопитающих он выполняет функции дыхания, питания, выделения, фильтрации и синтезе веществ, например гормонов.
|
|
|
Желточный мешок имеет энтодермальное происхождение, покрыт висцеральной мезодермой и непосредственно связан с кишечной трубкой зародыша. У зародышей с большим количеством желтка он принимает участие в питании. Желток не проходит через желточный проток, соединяющий мешок с кишкой. Сначала он переводится в растворимую форму под действием пищеварительных ферментов, продуцируемых энтодермальными клетками стенки мешка. Затем попадает в сосуды и с кровью разносится по всему телу зародыша.
У млекопитающих нет запасов желтка. Энтодерма желточного мешка у них служит местом образования первичных половых клеток, мезодерма дает форменные элементы крови зародыша. Кроме того, желточный мешок млекопитающих заполнен жидкостью, отличающейся высокой концентрацией аминокислот и глюкозы, что указывает на возможность обмена белков в желточном мешке.
Судьба желточного мешка у разных животных несколько различна. У птиц к концу периода инкубации остатки желточного мешка уже находятся внутри зародыша, после чего он быстро исчезает и к концу 6-х суток после вылупления полностью рассасывается. У млекопитающих желточный мешок бывает развит по-разному. У хищников он сравнительно большой, с сильно развитой сетью сосудов, а у приматов быстро сморщивается и исчезает без остатка до родов.
Аллантоис развивается несколько позднее других внезародышевых органов. Он представляет собой мешковидный вырост вентральной стенки задней кишки. Он образован энтодермой изнутри и спланхноплеврой снаружи. У рептилий и птиц аллантоис быстро дорастает до хориона и выполняет несколько функций. Прежде всего это вместилище для мочевины и мочевой кислоты, которые представляют собой конечные продукты обмена азотсодержащих органических веществ. В аллантоисе хорошо развита сосудистая сеть, благодаря чему вместе с хорионом он участвует в газообмене. При вылуплении наружная часть аллантоиса отбрасывается, а внутренняя — сохраняется в виде мочевого пузыря.
У многих млекопитающих аллантоис тоже хорошо развит и вместе с хорионом образует хориоаллантоисную плаценту (плацента означает тесное наложение или слияние зародышевых оболочек с тканями родительского организма).
По мере того как зародыш приобретает форму, вокруг него образуется несколько складок, которые как бы подсекают зародыш, отделяют его от желтка и устанавливают четкие границы между зародышем и внезародышевыми областями. Они называются туловищными складками. Сначала образуется головная складка. Она подсекает снизу головную часть. Задние концы этой складки переходят в боковые туловищные складки, отграничивающие туловище зародыша с боков. Хвостовая складка отграничивает задний конец зародыша. Постепенно сужается ножка, соединяющая среднюю кишку и желточный мешок, образуются передний и задний отделы кишки. Одновременно из эктодермы и прилежащей к ней соматоплевры образуется сначала головная складка, которая, как капюшон, нарастает на зародыш сверху. Концы головной складки образуют по бокам амниотические валики. Они растут поверх зародыша навстречу друг другу и срастаются, образуя сразу стенки амниона, прилежащего к зародышу, и хориона, лежащего снаружи.
|
|
|
63.Транскрипция и амплификация генов в овогенезе. Детерминация и процессы цитодифференцировки. Дифференциальная активность генов в эмбриогенезе. Образование органов и тканей. (Уверена только в образовании органов и тканей. Остальное не смогла найти нормально)
Транскрипция - переписывание информации с ДНК с целью ее использования для синтеза белка. Единицей транскрипции является транскриптон, включающий структурные и функциональные гены.
Амплификация генов- при этом крайне необычном процессе происходят серьёзные нарушения нормального процесса репликации ДНК. В результате вместо одной копии определённого участка хромосомы образуется множество копий. Это, в свою очередь, приводит к образованию множества копий генов, которые расположены в этом районе хромосомы. Иногда образуется так много копий амплифицированного региона, что они формируют собственные маленькие псевдохромосомы, которые называются «double-minute» хромосомами.

Органогенезы, заключающиеся в образовании отдельных органов, составляют основное содержание эмбрионального периода. Они продолжаются в личиночном и завершаются в ювенильном периоде. Необходимой предпосылкой перехода к органогенезам является достижение зародышем стадии гаструлы, а именно формирование зародышевых листков. Занимая определенное положение друг по отношению к другу, зародышевые листки, контактируя и взаимодействуя, обеспечивают такие взаимоотношения между различными клеточными группами, которые стимулируют их развитие в определенном направлении. Это так называемая эмбриональная индукция —важнейшее следствие взаимодействия между зародышевыми листками.
В ходе органогенезов изменяются форма, структура и химический состав клеток, обособляются клеточные группы, представляющие собой зачатки будущих органов. Постепенно развивается определенная форма органов, устанавливаются пространственные и функциональные связи между ними. Процессы морфогенеза сопровождаются дифференциацией тканей и клеток, а также избирательным и неравномерным ростом отдельных органов и частей организма. Обязательным условием органогенезов наряду с размножением, миграцией и сортировкой клеток является их избирательная гибель.
Самое начало органогенеза называют нейруляцией. Нейруляция охватывает процессы от появления первых признаков формирования нервной пластинки до замыкания ее в нервную трубку. Параллельно формируются хорда и вторичная кишка, а лежащая по бокам от хорды мезодерма расщепляется в краниокаудальном направлении на сегментированные парные структуры — сомиты.
Вначале неспециализированная спинная эктодерма, отвечая на индукционное воздействие со стороны хордомезодермы, превращается в нервную пластинку, представленную нейроэпителиальными клетками цилиндрической формы.

Рис. Последовательные стадии формирования нервной трубки и нервного гребня на 3-й неделе развития человеческого эмбриона (поперечный срез):
1—нервная пластинка, 2—нервный гребень, 3—эктодерма, 4—хорда, 5—нервная бороздка, 6—невроцель
Нервная пластинка недолго остается уплощенной. Вскоре ее боковые края приподнимаются, образуя нервные валики, которые лежат по обе стороны неглубокой продольной нервной бороздки. Края нервных валиков далее смыкаются, образуя замкнутую нервную трубку с каналом внутри — невроцелем. Раньше всего смыкание нервных валиков происходит на уровне начала спинного мозга, а затем распространяется в головном и хвостовом направлениях. После смыкания нервных валиков клетки, первоначально располагавшиеся между нервной пластинкой и будущей кожной эктодермой, образуют нервный гребень. Клетки нервного гребня отличаются способностью к обширным, но строго регулируемым миграциям по всему телу и образуют два главных потока. Клетки одного из них—поверхностного—включаются в эпидермис или дерму кожи, где дифференцируются в пигментные клетки. Другой поток мигрирует в брюшном направлении, образует чувствительные спинномозговые ганглии, симпатические нервные узлы, мозговое вещество надпочечников, парасимпатические ганглии. Клетки из черепного отдела нервного гребня дают начало как нервным клеткам, так и ряду других структур, таких, как жаберные хрящи, некоторые кроющие кости черепа.
Мезодерма, занимающая место по бокам от хорды и распространяющаяся далее между кожной эктодермой и энтодермой вторичной кишки, подразделяется на дорсальную и вентральную области. Дорсальная часть сегментирована и представлена парными сомитами. Закладка сомитов идет от головного к хвостовому концу. Вентральная часть мезодермы, имеющая вид тонкого слоя клеток, называется боковой пластинкой. Сомиты соединены с боковой пластинкой промежуточной мезодермой в виде сегментированных ножек сомитов.
Все области мезодермы постепенно дифференцируются. В начале формирования сомиты имеют конфигурацию, характерную для эпителия с полостью внутри. Под индукционным воздействием, исходящим от хорды и нервной трубки, вентромедиальные части сомитов — склеротомы —превращаются во вторичную мезенхиму, выселяются из сомита и окружают хорду и вентральную часть нервной трубки. В конце концов из них образуются позвонки, ребра и лопатки.
Дорсолатеральная часть сомитов с внутренней стороны образует миотомы, из которых разовьются поперечно-полосатые скелетные мышцы тела и конечностей. Наружная дорсолатеральная часть сомитов образует дерматомы, которые дают начало внутреннему слою кожи — дерме. Из области ножек сомитов с зачатками нефротом и гонотом образуются органы выделения и половые железы.
Правая и левая несегментированные боковые пластинки расщепляются на два листка, ограничивающих вторичную полость тела — целом. Внутренний листок, прилежащий к энтодерме, называют висцеральным. Он окружает кишку со всех сторон и образует брыжейку, покрывает легочную паренхиму и мышцу сердца. Наружный листок боковой пластинки прилежит к эктодерме и называется париетальным. В дальнейшем он образует наружные листки брюшины, плевры и перикарда.
Энтодерма у всех зародышей в конечном счете образует эпителий вторичной кишки и многие ее производные. Сама вторичная кишка всегда располагается под хордой.
Таким образом, в процессе нейруляции возникает комплекс осевых органов нервная трубка — хорда — кишка, представляющих собой характерную черту организации тела всех хордовых. Одинаковое происхождение, развитие и взаимное расположение осевых органов выявляют их полную гомологию и эволюционную преемственность.
При углубленном рассмотрении и сравнении процессов нейруляции у конкретных представителей типа хордовых выявляются некоторые различия, которые связаны в основном с особенностями, зависящими от строения яйцеклеток, способа дробления и гаструляци. Обращают внимание отличающаяся форма зародышей и смещение времени закладки осевых органов друг относительно друга, т.е. описанная выше гетерохрония.
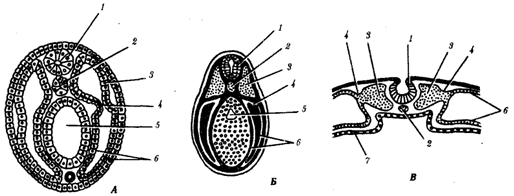
Рис. Нейрулы различных хордовых животных.
А — ланцетник; Б —лягушка; В — цыпленок:
1—нервная трубка, 2—хорда, 3—сомит, 4—ножка сомита, 5—вторичная кишка, 6—боковая пластинка, 7—энтодерма
Эктодерма, мезодерма и энтодерма в ходе дальнейшего развития, взаимодействуя друг с другом, участвуют в формировании определенных органов. Возникновение зачатка органа связано с местными изменениями определенного участка соответствующего зародышевого листка. Так, из эктодермы развиваются эпидермис кожи и его производные (перо, волосы, ногти, кожные и молочные железы), компоненты органов зрения; слуха, обоняния, эпителий ротовой полости, эмаль зубов. Важнейшими эктодермальными производными являются нервная трубка, нервный гребень и образующиеся из них все нервные клетки.
Производными энтодермы являются эпителий желудка и кишки, клетки печени, секретирующие клетки поджелудочной, кишечных и желудочных желез. Передний отдел эмбриональной кишки образует эпителий легких и воздухоносных путей, а также секретирующие клетки передней и средней долей гипофиза, щитовидной и паращитовидной желез.
Мезодерма помимо уже описанных выше скелетных структур, скелетной мускулатуры, дермы кожи, органов выделительной и половой систем образует сердечно-сосудистую систему, лимфатическую систему, плевру, брюшину и перикард. Из мезенхимы, имеющей смешанное происхождение за счет клеток трех зародышевых листков, развиваются все виды соединительной ткани, гладкая мускулатура, кровь и лимфа.
Зачаток конкретного органа формируется первоначально из определенного зародышевого листка, но затем орган усложняется и в итоге в его формировании принимают участие два или три зародышевых листка.
64. Опыты экспериментальной эмбриологии. Эмбриональная индукция как процесс взаимодействия между частями развивающегося зародыша. Индукторы и индукционные взаимодействия в эмбриогенезе. Каскадная индукция. Примеры. ( Получилось так, что вся глава из книги здесь. Не знаю, что убрать ).
Экспериментальная эмбриология - раздел эмбриологии, изучающий причинные механизмы индивидуального развития животных и растений при помощи экспериментов на живых организмах, использующий методы маркировки, удаления, пересадки и изоляции частей тела и органов, а также различные воздействия внешними факторами. Э.Э. выясняет стадии детерминации материала зачатков органов и тканей, источники формативных или индукционных влияний, значение синтеза макромолекул в процессах детерминации и дифференцировки, факторы формообразования. С помощью удаления, инактивации и пересадки клеточных ядер изучают взаимодействия ядра и цитоплазмы в периоды гаметогенеза и зародышевого развития, стадии и факторы дифференциальной активации генов в процессе развития и другие проблемы.
Эмбриональная индукция — это взаимодействие частей развивающегося зародыша, при котором один участок зародыша влияет на судьбу другого участка. Явление эмбриональной индукции с начала XX в. изучает экспериментальная эмбриология.
Классическими считают опыты немецкого ученого Г. Шпемана и его сотрудников (1924) на зародышах амфибий. Для того чтобы иметь возможность проследить за судьбой клеток определенного участка зародыша, Шпеман использовал два вида тритонов: тритона гребенчатого, яйца которого лишены пигмента и потому имеют белый цвет, и тритона полосатого, яйца которого благодаря пигменту имеют желто-серый цвет.
Один из опытов заключается в следующем: кусочек зародыша из области дорсальной губы бластопора на стадии гаструлы тритона гребенчатого пересаживают на боковую или вентральную сторону гаструлы тритона полосатого (рис. 8.8). В месте пересадки происходит развитие нервной трубки, хорды и других органов. Развитие может достичь довольно продвинутых стадий с образованием дополнительного зародыша на боковой или вентральной стороне зародыша реципиента. Дополнительный зародыш содержит в основном клетки зародыша реципиента, но светлые клетки зародыша-донора тоже обнаруживаются в составе различных органов.

Рис. 8.8. Пересадка спинной губы от зародыша-донора на брюшную сторону зародыша-реципиента. А—схема опыта; Б—поперечный срез на стадии закладки двух комплексов осевых органов:
1—первичный зародыш, 2—вторичный, индуцированный зародыш
Из этого и подобных опытов следует несколько выводов. Во-первых, участок, взятый из спинной губы бластопора, способен направлять или даже переключать развитие того материала, который находится вокруг него, на определенный путь развития. Он как бы организует, или индуцирует, развитие зародыша как в обычном, так и в нетипичном месте. Во-вторых, боковая и брюшная стороны гаструлы обладают более широкими потенциями к развитию, нежели их презумптивное (предполагаемое) проспективное направление, так как вместо обычной поверхности тела в условиях эксперимента там образуется целый зародыш. В-третьих, достаточно точное строение новообразованных органов в месте пересадки указывает на эмбриональную регуляцию. Это означает, что фактор целостности организма приводит к достижению хорошего конечного результата из нетипичных клеток в нетипичном месте, как бы управляя процессом, регулируя его в целях достижения этого результата.
Г. Шпеман назвал спинную губу бластопора первичным эмбриональным организатором. Первичным потому, что на более ранних стадиях развития подобных влияний обнаружить не удавалось, а организатором потому, что влияние происходило именно на морфогенез. В настоящее время установлено, что главная роль в спинной губе бластопора принадлежит хордомезодермальному зачатку, который назвали первичным эмбриональным индуктором, а само явление, при котором один участок зародыша влияет на судьбы другого,—эмбриональной индукцией.
В 30-е гг. исследователи пытались установить природу индуцирующего действия. Вскоре выяснилось, что разнообразные убитые ткани, вытяжки из самых различных тканей беспозвоночных и позвоночных животных, а также растений, несколько классов химических соединений (белки, нуклеопротеины, стероиды и даже неорганические вещества) могут вызывать индукцию. Таким образом, была установлена химическая природа организаторов. Одновременно стало ясно, что специфичность ответа прямо не связана с химическими свойствами индуктора.
Внимание эмбриологов переключилось на индуцируемые ткани. Оказалось, что специфичность действия индуктора-раздражителя может быть весьма различной, а сам эффект индуцирующего воздействия ограничивается способностью того или иного участка развивающегося зародыша воспринимать это воздействие и отвечать на него.
Некоторые индукторы, по-видимому, более или менее специфичны в определении судьбы индуцируемой ткани. Об этом свидетельствуют следующие опыты. Если пересадить спинную губу ранней гаструлы, то индуцируется развитие структур переднего мозга (головной индуктор), если же пересадить спинную губу поздней гаструлы, то развиваются спинной мозг и мезодермальные ткани. Было показано также, что наиболее сильное нейрализующее влияние оказывает фракция нуклеопротеинов, а мезодермализующим индуктором оказался белок. Если имплантировать оба эти индуктора в виде смеси клеток или смеси веществ, то получаются хорошо развитые зародыши.

Рис. 8.9. Результаты пересадки головного (А) и туловищного (Б) индукторов
Другие индукторы действуют как неспецифические пусковые механизмы, как бы высвобождая ответ, уже детерминированный в клетках индуцируемой ткани. Было показано, что, например, слуховой пузырек выступает не только в роли индуктора слухового аппарата, но и является активатором различных морфогенетических процессов. Будучи пересажен в область боковой линии эмбриона тритона, он влечет за собой индукцию конечности. Конечность можно индуцировать также пересадкой носовой плакоды или гипофиза. Легче всего добавочные конечности индуцируются в области боковой линии, но они могут быть получены и на брюшной стороне. Эти примеры указывают на то, что специфический ответ зависит не столько от индуктора, сколько отреагирующей области.
Способность эмбрионального материала реагировать на различного рода влияния изменением своей презумптивной судьбы получила название компетенции. Установлено, например, что компетенция к образованию нервной системы у амфибий затрагивает всю эмбриональную эктодерму и возникает с момента начала гаструляции. К концу гаструляции эта компетенция прекращается. Таким образом, изменение хода развития возможно лишь в том случае, если область компетенции к образованию некоторой закладки шире, чем область, из которой она в норме развивается, а также если индукционное действие происходит в определенный интервал онтогенетического развития.
Изучение индукционных взаимодействий у разных представителей типа хордовых показало, что области и сроки компетенции неодинаковы. Так, у асцидий на стадии 8 бластомеров, когда уже все основные зачатки предопределены, проводили некоторые перемещения бластомеров. Материал хордомезодермы и основная часть нейрального материала у них локализованы в заднем вегетативном бластомере. Небольшая часть нейрального материала, формирующего головной ганглий, находится в заднем анимальном бластомере, расположенном над задним вегетативным (рис. 8.10).
Для проверки индукционных взаимодействий между ними анимальный ярус бластомеров поворачивали на 180° так, чтобы задний анимальный бластомер терял контакт с задним вегетативным. Головной ганглий не развился нигде. Это означает, что для развития головного ганглия необходимо индукционное влияние на задний анимальный бластомер со стороны заднего вегетативного. Кроме того, очевидно, что задний анимальный бластомер не обладает автономностью развития, но только он компетентен к восприятию воздействия со стороны заднего вегетативного бластомера, содержащего хордомезодермальный зачаток.
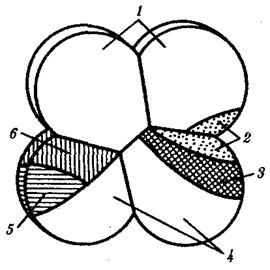
Рис. 8.10. Карта презумптивных зачатков у зародыша асцидий
на стадии восьми бластомеров:
1—эпидермис, 2—нервная пластинка, 3—хорда, 4—энтодерма, 5—сомиты, 6—мезенхима
Во всех других классах хордовых индукционные взаимодействия между хордомезодермальным и нейральным зачатками подобны таковым у амфибий. Полагают, что в ходе эволюции хордовых произошли расширение областей и удлинение срока компетенции. Это расценивают как признак существенного эволюционного прогресса.
Явления индукции многочисленны и разнообразны. Помимо первичной индукции со стороны спинной губы бластопора описаны индукционные влияния на более поздних, нежели гаструляция, этапах развития. Все они являются вторичными и третичными, представляя собой каскадные взаимодействия, типичные для дифференцировки, потому что индукция многих структур зависит от предшествующих индукционных событий. Примером вторичной индукции может служить действие глазного бокала (выпячивание переднего мозга) на прилежащий покровный эпителий, под влиянием чего эпителий впячивается, а затем отшнуровывается хрусталиковый пузырек—зачаток глазного хрусталика (рис. 8.11). Расположенный над хрусталиком покровный эпителий тоже испытывает сложные изменения, теряет пигмент и становится роговичным эпителием. Это пример третичной индукции. Таким образом получается, что глазной бокал возникает только после развития передней части головного мозга, хрусталик — после формирования бокала, а роговица — после образования хрусталика.
Вместе с тем индукция носит не только каскадный, но и переплетающийся характер, т.е. в индукции той или иной структуры может участвовать не одна, а несколько тканей. В свою очередь, такая структура может служить индуктором для нескольких других тканей. Например, глазной бокал служит главным, но не единственным индуктором хрусталика. Морфогенез всегда сопровождается значительными перемещениями тканей друг относительно друга. Так, презумптивный хрусталик, т.е. эпидермис, из которого в последующем должен развиться хрусталик, во время гаструляции лежит над энтодермой будущей глотки, которая служит первым индуктором хрусталика. Затем под этим эпидермисом оказывается сердечная мезодерма, которая тоже действует как индуктор. И только позднее, во время нейруляции на переднем конце нервной трубки выпячиваются глазные пузыри, образующие глазной бокал и сетчатку, являющуюся главным индуктором хрусталика (рис. 8.12).

Рис. 8.11. Развитие (А —Г) глаза у хвостатой амфибии:
1—хрусталиковая плакода, 2—пигментный эпителий, 3—сетчатка, 4—роговица, 5—хрусталиковые волокна, 6—хрусталиковый эпителий, 7—глазной бокал

Рис. 8.12. Последовательные индукционные взаимодействия,
необходимые для образования хрусталика у зародыша амфибии:
I—ранний зародыш, II—гаструла, III—нейрула, IV—стадия хвостовой почки, V—личинка, VI—взрослая особь; 1—плакода, 2—пузырек, 3—волокна, а—энтодерма, б—сердечная мезодерма, в—сетчатка
Удаляя ту или иную из индуцирующих тканей, определили степень участия каждой из них в индукции хрусталика. Оказалось, что при удалении сетчатки глазного бокала у 42% зародышей амфибий все же формировались хрусталики и, следовательно, энтодерма и мезодерма в сумме обладают почти таким же индуцирующим действием, как и сетчатка глазного бокала. Полагают, что многочисленность индуцирующих тканей может иметь решающее значение для точного установления места формирования органа. Кроме того, сети индукции могут играть важную роль в канализации развития, обеспечивая нормальное течение органогенеза, даже если один из компонентов ин-
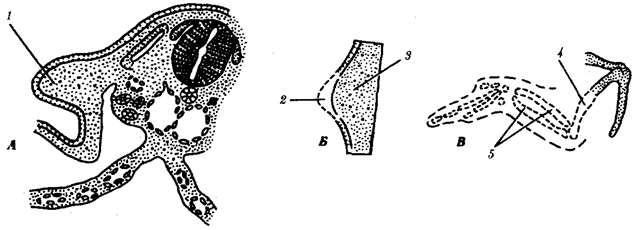
Рис. 8.13. Влияние удаления апикального эктодермального гребня
на развитие почки крыла.
А—схема расположения почки крыла; Б—почка крыла; В—недоразвитие скелета (пунктир) после удаления апикального гребня:
1—почка крыла, 2 —апикальный эктодермальный гребень, 3—мезенхима, 4 —плечевая кость, 5—локтевая и лучевая кости
Чаще всего близлежащие участки зародыша оказывают взаимное влияние друг на друга. Демонстративным примером являются взаимодействия в зачатке конечности. Конечность развивается из скопления клеток, происходящих из боковой мезодермы, и покрывающих их клеток эктодермы (рис. 8.13). Развитие конечности начинается с активации клеток боковой мезодермы в непосредственной близости от сомитов, которые, возможно, и оказывают индуцирующие импульсы на мезодерму в области будущей конечности. Активированные мезодермальные клетки зачатка конечности влияют на покрывающую их эктодерму, в результате чего она утолщается. Образовавшееся утолщение эпидермиса на его верхушке называют апикальным эктодермальным гребнем. Последний стимулирует рост почки конечности (при удалении его рост почки конечности прекращается). Мезодерма же поддерживает гребень в активном состоянии и определяет форму конечности. Например, мезодерма из почки крыла при соединении с эктодермой почки ноги образует крыло, покрытое перьями, или мезодерма из почки конечности утиного зародыша с эктодермой куриного приводит к развитию перепончатой конечности.
Различают гетерономную и гомономную виды индукции. К гетерономной относят случаи, подобные описанному, при которых один кусочек зародыша индуцирует иной орган (хордомезодерма индуцирует появление нервной трубки и всего зародыша в целом).Гомономная индукция заключается в том, что индуктор побуждает окружающий материал к развитию в том же направлении, что и он сам. Например, область нефротома, пересаженная другому зародышу, способствует развитию окружающего материала в сторону формирования головной почки, а прибавление в культуру фибробластов сердца маленького кусочка хряща влечет за собой процесс образования хряща.
Чтобы воспринять действие индуктора, компетентная ткань должна обладать хотя бы минимальной организацией. Одиночные клетки не воспринимают действие индуктора, а чем больше клеток в реагирующей ткани, тем активнее ее реакция. Для оказания индуцирующего действия иногда достаточно лишь одной клетки индуктора.
Индукционные взаимодействия могут проявляться в культуре ткани in vitro, но по-настоящему полноценными они бывают только в структуре целостного организма.
Весьма интересны результаты опытов, помогающие оценить взаимосвязь индукционных взаимодействий с цитодифференцировкой и морфогенезом. Ранее уже было описано определяющее влияние мезенхимы на морфогенез конечностей позвоночных.
Многочисленными опытами показано также большое влияние мезенхимы на морфогенез желез эпителиального происхождения. Легочная энтодерма, например, при выращивании с печеночной мезенхимой приобретает строение печеночных балок, а эпителий молочной железы под влиянием мезенхимного зачатка слюнной железы приобретает морфологию слюнной железы. Это происходит как при выращивании in vitro, так и при трансплантации в организм животного-реципиента. Подобные результаты с несомненностью указывают на необходимость индуцирующего влияния мезенхимы на морфогенез.
Однако не менее интересен факт, что морфогенез не всегда сопряжен с определенным направлением дифференцирован эпителия. Так, рекомбинантная слюнная железа, полученная из зачатка молочной железы и мезенхимы слюнной, при подсадке лактирующей самке-реципиенту начинает вырабатывать молоко несмотря
|
|
|
