 |


Биосинтез пиримидиновых рибонуклеотидов и его регуляция. Оротацидурия. «Запасные» пути синтеза пиримидиновых оснований.
|
|
|
|
Синтез пуриновых нуклеотидов, регуляция. Образование 5-фосфорибозиламина, происхождение атомов пуринового ядра, ИМФ как общий предшественник АМФ и ГМФ, регуляция. «Запасные пути» синтеза пуриновых нуклеотидов.
II. СИНТЕЗ ПУРИНОВЫХ НУКЛЕОТИДОВ
В 40-50-х годах XX столетия опытами с мечеными изотопами удалось выяснить происхождение атомов пуринового ядра при синтезе пуринов de novo. Было установлено, что в формировании кольца принимают участие аминокислоты Асп, Гли, Глн, СО2 и два одноуглеродных производных тетрагидрофолата: метенил-Н4-фолат и формил-Н4-фолат. Этим способом образуется основное количество пуриновых нуклеотидов, тогда как нуклеотиды, синтезирующиеся за счёт повторного использования азотистых оснований или нуклеозидов, составляют не более 10-20% общего фонда этих соединений.
А. Образование 5-фосфорибозил-1-дифосфата
Фосфорибозилдифосфат (ФРДФ), или фос-форибозилпирофосфат (ФРПФ) занимает центральное место в синтезе как пуриновых, так и пиримидиновых нуклеотидов (рис. 10-2).
Он образуется за счёт переноса β,γ-пирофосфатного остатка АТФ на рибозо-5-фосфат в реакции, катализируемой ФРДФ-синтетазой.
Источниками рибозо-5-фосфата могут быть: пентозофосфатный путь превращения глюкозы или катаболизм нуклеозидов, в ходе которого под действием нуклеозидфосфорилазы первоначально образуется рибозо-1-фосфат, а затем с помощью соответствующей мутазы фосфатный остаток переносится в 5-положение.
ФРДФ участвует не только в синтезе пуриновых и пиримидиновых нуклеотидов из простых предшественников (т.е. de novo), но используется на образование пуриновых нуклеотидов по "запасному" пути и в синтезе нуклеотидных коферментов.
|
|
|
Б. Биосинтез пуриновых нуклеотидов DE NOVO
Сборка пуринового гетероцикла осуществляется на остатке рибозо-5-фосфата при участии различных доноров углерода и азота (рис. 10-3).

Рис. 10-1. Переваривание нуклеиновых кислот пищи.
Включение простых предшественников в нуриновое кольцо с образованием ИМФ
Первая специфическая реакция образования пуриновых нуклеотидов - перенос амидной группы Глн на ФРДФ с образованием 5-фосфорибозил-1 -амина (рис. 10-4). Эту реакцию катализирует фермент амидофосфорибозилтрансфераза. При этом формируется β-N-гликозидная связь.
Затем к аминогруппе 5-фосфорибозил-1-амина присоединяются остаток глицина, N5,N10-метенил-Н4-фолата ещё одна амидная группа глутамина, диоксид углерода, аминогруппа аспартата и формильный остаток N10-формил Н4-фолата. Результатом этой десятистадийной серии реакций является образование первого пуринового нуклеотида - инозин-5'-монофосфата (ИМФ), на синтез которого затрачивается не менее шести молекул АТФ. В отличие от прокариотов, у которых каждую стадию этого процесса катализирует отдельный фермент, у эукариотов за счёт слияния генов возникли полифункциональные ферменты, каждый из которых катализирует несколько реакций. В синтезе пуриновых нуклеотидов de novo это реакции 3,4 и 6, 7-8 и 10-11 соответственно.

Рис. 10-2. Образование 5-фосфорибозил-1-дифосфата.

Рис. 10-3. Происхождение атомов С и N в пуриновом кольце.
ИМФ в основном используется на синтез АМФ или ГМФ. Небольшое количество этого продукта обнаруживается также в тРНК в качестве одного из минорных нуклеотидов.
Превращение ИМФ в АМФ и ГМФ в обоих случаях включает 2 стадии и идёт с затратой энергии (рис. 10-5).
Аденилосукцинатсинтетаза, используя энергию ГТФ, присоединяет аспартат к ИМФ с образованием аденилосукцината, который в реакции, катализируемой аденилосукциназой, отщепляет фумарат и превращается в АМФ.
|
|
|
Второй пуриновый нуклеотид (ГМФ) образуется также в 2 стадии. Сначала ИМФ окисляется NAD+-зависимой ИМФ-дегидрогеназой с образованием ксантозин-5'-монофосфата (КМФ). Последующее трансамидирование гидроксильной группы при С2-пуринового кольца КМФ катализирует ГМФ-синтетаза с использованием амидной группы Глн и энергии АТФ.
При образовании пуриновых нуклеотидов ГТФ расходуется на синтез АМФ, а АТФ - на синтез ГМФ. Перекрёстное использование пуриновых нуклеозидтрифосфатов на образование конечных продуктов синтеза помогает поддерживать в клетках баланс адениловых и гуаниловых нуклеотидов.
Печень - основное место образования пури-новых нуклеотидов, откуда они могут поступать в ткани, не способные к их синтезу: эритроциты, ПЯЛ и частично мозг.
Образование нуклеозид- ди- и трифосфатов
В образовании нуклеиновых кислот, некоторых коферментов и во многих синтетических процессах нуклеотиды используются в виде ди-и трифосфатов, синтез которых катализируют ферменты класса трансфераз. АМФ и ГМФ превращаются в нуклеозиддифосфаты (НДФ) с помощью специфичных к азотистому основанию нуклеозидмонофосфаткиназ (НМФ-киназ) и АТФ. Так, аденилаткиназа катализирует реакцию:
АМФ + АТФ → 2 АДФ,
а гуанилаткиназа:
ГМФ + АТФ → ГДФ + АДФ.
Аденилаткиназа особенно активна в печени и мышцах, где высок уровень энергоёмких процессов. Функция этого фермента заключается в том, чтобы поддерживать в тканях равновесие фонда адениловых нуклеотидов: АМФ, АДФ и АТФ.
Взаимопревращения нуклеозиддифосфатов и нуклеозидтрифосфатов осуществляет нуклеозид-дифосфаткиназа. Этот фермент в отличие от НМФ-киназ обладает широкой субстратной специфичностью и, в частности, может катализировать реакцию:
ГДФ + АТФ → ГТФ + АДФ.
Превращение АДФ в АТФ происходит, в основном, за счёт окислительного фосфорилирования или в реакциях субстратного фосфорилирования гликолиза или цитратного цикла.
В. "Запасные" пути синтеза путиловых нуклеотидов (реутилизация азотистых оснований и нуклеозидов)
Огромные затраты энергии для синтеза пуриновых нуклеотидов de novo не способны полностью обеспечить субстратами синтез нуклеиновых кислот в период гаструляции и раннего роста ребёнка. Потребность в большом количестве нуклеотидов привела к развитию "запасных" путей синтеза этих "дорогих" молекул. Наибольшее значение в этом процессе имеют ферменты, осуществляющие превращение пуринов в мононуклеотиды с использованием ФРДФ как донора остатка фосфорибозы.
|
|
|
Синтез АМФ и ГМФ из аденина и гуанина
ФРДФ-зависимое фосфорибозилирование пуринов катализируют 2 фермента.
Аденинфосфорибозилтрансфераза, ответственная за образование АМФ (рис. 10-6).
Гипоксантин-гуанинфосфорибозилтрансфераза, катализирующая образование ИМФ и ГМФ из гипоксантина и гуанина соответственно (рис. 10-7).
Однако в организме при любых ситуациях этот путь синтеза пуриновых нуклеотидов, получивший название "путь спасения", имеет вспомогательное значение.
Нуклеозидкиназы
Нуклеозиды, получающиеся при катаболизме нуклеиновых кислот из нуклеотидов под действием нуклеотидаз, могут повторно фосфорилироваться, образуя нуклеозид-5'-монофосфаты за счёт переноса γ-фосфатного остатка АТФ на соответствующий субстрат. У млекопитающих такой путь пополнения запасов пуриновых нуклеотидов в клетке не имеет существенного значения. Основным ферментом этой группы является аденозинкиназа, которая ускоряет реакцию:
Аденозин + АТФ → АМФ + АДФ.
Из всех способов реутилизации пуринов наиболее активна гипоксантин-гуанинфосфорибозилтрансферазная реакция, поскольку ИМФ, образующийся в этой реакции, вовлекается в синтез АМФ и ГМФ. Использование гипоксантина и гуанина по запасному пути становится жизненно важным событием в клетках, не способных к синтезу пуриновых нуклеотидов de novo. Значение аденинфосфорибозилтрансферазы в повторном использовании аденина менее существенно. По сравнению с аденозином количество аденина в клетках мало, а первый возвращается в фонд нуклеотидов с помощью аденозинкиназы.

Рис. 10-6. Фосфорибозилирование аденина в АМФ.
Рис. 10-7. Фосфорибозилирование гипоксантина и гуанина с образованием ИМФ и ГМФ.
Г. Регуляция синтеза пуриновых нуклеотидов
|
|
|
Основным показателем, от которого зависит синтез пуриновых нуклеотидов, служит концентрация ФРДФ, которая, в свою очередь, зависит от скорости его синтеза, утилизации и разрушения. Количество ФРДФ определяется доступностью рибозо-5-фосфата и активностью ФРДФ синтетазы - фермента, чувствительного к концентрации фосфата и пуриновых нуклеотидов. Внутриклеточная концентрация ФРДФ строго регулируется и обычно низкая. ФРДФ синтетаза - аллостерический фермент. Он активируется неорганическим фосфатом (Рi) и ингибируется пуриновыми нуклеозид- моно-, ди- и трифосфатами, которые по эффективности ингибирования распределяются в следующем порядке:
НМФ > НДФ > НТФ (рис. 10-8). ФРДФ служит не только субстратом, но и аллостерическим активатором второй реакции синтеза пуринонуклеотидов de novo, которую катализирует амидофосфорибозилтрансфераза.

Рис. 10-8. Регуляция синтеза пуриновых нуклеотидов. 1 - ФРДФ синтетаза; 2 - амидофоофорибозилтрансфераза; 3 - ИМФ дегидрогеназа; 4 - аденилосукцинатсинтетаза.
Пуриновые нуклеотиды, особенно АМФ и ГМФ по механизму отрицательной обратной св1язи ингабируют амидофосфорибозилтрансферазу, которая катализирует первую специфическую реакцию синтеза пуриновых нуклеотидов de novo.
Метаболическая цепь образования АМФ и ГМФ de novo регулируется также в месте её разветвления: АМФ ингибирует аденилосукцинатсинтетазу, а ГМФ - реакцию образования ксантиловой кислоты, которую катализирует ИМФ дегидрогеназа. Перекрёстная регуляция путей использования ИМФ служит для того, чтобы снизить синтез одного пуринового нуклеотида при дефиците другого.
Помимо ферментов основного пути синтеза пуриновых нуклеотидов de novo, регулируется также активность ферментов "запасных" путей: аденинфосфорибозилтрансфераза ингибируется АМФ, а гипоксантин-гуанинфосфорибозилтрансфераза - ИМФ и ГМФ.
63. Катаболизм пуриновых и пиримидиновых нуклеотидов: основные этапы, конечные продукты. Причины и проявления гиперурикемии; подагра, синдром Леша-Нихена. Применение аллопуринола для лечения подагры
III. КАТАБОЛИЗМ ПУРИНОВЫХ НУКЛЕОТИДОВ
У человека основной продукт катаболизма пуриновых нуклеотидов - мочевая кислота. Её образование идёт путём гидролитического отщепления фосфатного остатка от нуклеотидов с помощью нуклеотидаз или фосфатаз, фосфоролиза N-гликозидной связи нуклеозидов пуриннуклеозидфосфорилазой, последующего дезами-нирования и окисления азотистых оснований (рис. 10-9).
От АМФ и аденозина аминогруппа удаляется гидролитически аденозиндезаминазой с образованием ИМФ или инозина. ИМФ и ГМФ превращаются в соответствующие нуклеозиды: инозин и гуанозин под действием 5´-нуклеотидазы. Пуриннуклеозидфосфорилаза катализирует расщепление N-гликозидной связи в инозине и гуанозине с образованием рибозо-1-фосфата и азотистых оснований: гуанина и гипоксантина. Гуанин дезаминируется и превращается в ксантин, а гипоксантин окисляется в ксантин с помощью ксантиноксидазы, которая катализирует и дальнейшее окисление ксантина в мочевую кислоту.
|
|
|
Ксантиноксидаза - аэробная оксидоредуктаза, простетическая группа которой включает ион молибдена, железа (Fe3+) и FAD. Подобно другим оксидазам, она окисляет пурины молекулярным кислородом с образованием пероксида водорода. В значительных количествах фермент обнаруживается только в печени и кишечнике.
Мочевая кислота удаляется из организма главным образом с мочой и немного через кишечник с фекалиями. У всех млекопитающих, кроме приматов и человека, имеется фермент уриказа, расщепляющий мочевую кислоту с образованием аллантоина, хорошо растворимого в воде (рис. 10-10).
Амфибии, птицы и рептилии, подобно человеку, лишены уриказы и экскретируют мочевую

Рис. 10-9. Катаболизм пуриновых нуклеотидов до мочевой кислоты.
кислоту и гуанин в качестве конечных продуктов обмена.
Мочевая кислота является слабой кислотой. Содержание недиссоциированной формы и солей (уратов) зависит от рН раствора. При физиологических значениях рН у мочевой кислоты может диссоциировать только один протон из трёх (рК = 5,8), поэтому в биологических жидкостях присутствует как недиссоциированная кислота в комплексе с белками, так и её натриевая соль.
В сыворотке крови в норме содержание мочевой кислоты составляет 0,15-0,47 ммоль/л или 3-7 мг/дл. Ежесуточно из организма выводится от 0,4 до 0,6 г мочевой кислоты и уратов.
IV. НАРУШЕНИЯ ОБМЕНА ПУРИНОВЫХ НУКЛЕОТИДОВ
Ураты значительно более растворимы, чем мочевая кислота: так, в моче с рН 5,0, когда

Аллантоин Рис. 10-10. Превращение мочевой кислоты в аллантоин.
мочевая кислота не диссоциирована, ее растворимость в 10 раз меньше, чем в моче с рН 7,0, при котором основная часть мочевой кислоты представлена солями. Реакция мочи зависит от состава пищи, но, как правило, она слабокислая, поэтому большинство камней в мочевыводящей системе - кристаллы мочевой кислоты.
А. Гиперурикемия подагры
Когда в плазме крови концентрация мочевой кислоты превышает норму, то возникает гиперурикемия. Вследствие гиперурикемии может развиться подагра - заболевание, при котором кристаллы мочевой кислоты и уратов откладываются в суставных хрящах, синовиальной оболочке, подкожной клетчатке с образованием подагрических узлов, или тофусов. К характерным признакам подагры относят повторяющиеся приступы острого воспаления суставов (чаще всего мелких) - так называемого острого подагрического артрита. Заболевание может прогрессировать в хронический подагрический артрит.
Поскольку лейкоциты фагоцитируют кристаллы уратов, то причиной воспаления является разрушение лизосомальных мембран лейкоцитов кристаллами мочевой кислоты. Освободившиеся лизосомальные ферменты выходят в иитозоль и разрушают клетки, а продукты клеточного катаболизма вызывают воспаление.
Общий фонд сывороточных уратов в норме составляет ~ 1,2 г у мужчин и 0,6 г у женщин. При подагре без образования тофусов (т.е. подагрических узлов, в которых накапливаются ураты натрия и мочевая кислота) количество уратов возрастает до 2-4 г, а у пациентов с тяжёлой формой болезни, сопровождающейся ростом тофусов, может достигать 30 г.
Подагра - распространённое заболевание, в разных странах ею страдают от 0,3 до 1,7% населения. А поскольку сывороточный фонд уратов у мужчин в 2 раза больше, чем у женщин, то они и болеют в 20 раз чаще, чем женщины.
Как правило, подагра генетически детерминирована и носит семейный характер. Она вызвана нарушениями в работе ФРДФ синтетазы или ферментов "запасного" пути: гипоксантин-гуанин- или аденинфосфорибозилтрансфераз.
К другим характерным проявлениям подагры относят нефропатию, при которой наблюдают образование уратных камней в мочевыводящих путях.
Полиморфные варианты ФРДФ синтетазы
Активность ФРДФ синтетазы, катализирующей образование ФРДФ, строго контролируется пуриновыми нуклеотидами. Мутации в гене ФРДФ синтетазы привели к появлению полиморфных вариантов фермента, которые характеризуются аномальным ответом на обычные регуляторные факторы: концентрацию рибозо-5-фосфата и пуриннуклеотидов. Как правило, наблюдается суперактивация фермента. Пуриновые нуклеотиды синтезируются со скоростью, почти независимой от нужд клетки. Это вызывает ингибирование запасных "путей спасения", усиление катаболизма избыточного количества нуклеотидов, повышение продукции мочевой кислоты, гиперурикемию и подагру (табл. 10-1).
Примерно у 40% больных одной из форм гликогеноза - болезнью Гирке (недостаточностью глюкозо-6-фосфатазы) сопутствующей патологией является подагра. Снижение способности печени секретировать глюкозу в кровь увеличивает использование глюкозо-6-фосфата в пентозофосфатном пути. Образуются большие количества рибозо-5-фосфата, которые могут стимулировать избыточный синтез, а следовательно, и катаболизм пуриновых нуклеотидов.
Б. Недостаточность ферментов "запасных путей" синтеза пуриновых нуклеотидов. Синдром Лёша-Нихена
В ряде случаев причиной гаперурикемии, избыточной экскреции пуринов с мочой и подагры являются нарушения в работе ферментов "пути спасения" пуриновых оснований (табл. 10-1). Гипоксантин-гуанин фосфорйбозилтранс-фераза катализирует реакцию превращения гуанина и гапоксантина в соответствующие нукле-отиды (рис. 10-7). Обнаружены полиморфные варианты гипоксантин-гуанинфосфорибозил-трансферазы со сниженной ферментативной активностью, что:
уменьшает повторное использование пуриновых оснований, и они превращаются в мочевую кислоту;
увеличивает синтез пуриновых нуклеотидов de novo из-за слабого использования ФРДФ в реакциях реутилизации и увеличения его концентрации в клетке. Адениловые и гуаниловые нуклеотиды образуются в количествах, превышающих потребности клеток, а это способствует усилению их катаболизма.
Синдром Лёша-Нихена - тяжёлая форма гиперурикемии, которая наследуется как рецессивный признак, сцепленный с Х-хромосомой, и проявляется только у мальчиков.
Болезнь вызвана полным отсутствием активности гипоксантин-гуанинфоефорибозилтранс-феразы и сопровождается гиперурикемией с содержанием мочевой кислоты от 9 до 12 мг/дл, что превышает растворимость уратов при нормальном рН плазмы. Экскреция мочевой кислоты у больных с синдромом Лёша-Нихена превышает 600 мг/сут и требует для выведения этого количества продукта не менее 2700 мл мочи.
У детей с данной патологией в раннем возрасте появляются тофусы, уратные камни в моче-выводящих путях и серьёзные неврологические отклонения, сопровождающиеся нарушением речи, церебральными параличами, снижением интеллекта, склонностью к нанесению себе увечий (укусы губ, языка, пальцев).
В первые месяцы жизни неврологические расстройства не обнаруживаются, но на пелёнках отмечают розовые и оранжевые пятна, вызванные присутствием в моче кристаллов мочевой кислоты. При отсутствии лечения больные погибают в возрасте до 10 лет из-за нарушения функции почек.
Полная потеря активности аденинфосфорибозилтрансферазы не столь драматична, как отсутствие
Таблица 10-1. Гиперурикемия, вызванная дефектами в работе ферментов обмена пуриннуклеотидов 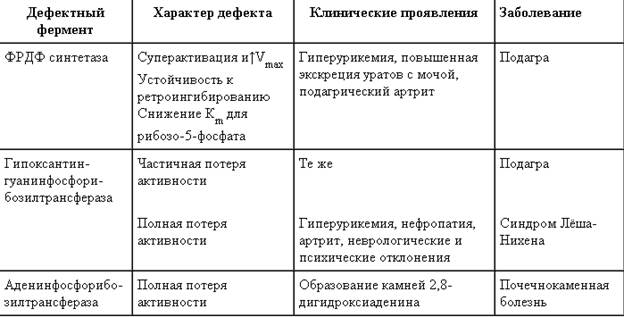
гипоксантин-гуанинфосфорибозилгрансферазы, однако и в этом случае нарушение повторного использования аденина вызывает гиперурикемию и почечнокаменную болезнь, при которой наблюдается образование кристаллов 2,8-дигидроксиаденина.
В. Лечение гиперурикемии
Основным препаратом, используемым для лечения гиперурикемии, является аллопуринол - структурный аналог гипоксантина (рис. 10-11).

Рис. 10-11. Строение аллопуринола и гипоксантина.
Аллопуринол оказывает двоякое действие на обмен пуриновых нуклеотидов:
ингибирует ксантиноксидазу и останавливает катаболизм пуринов на стадии образования гипоксантина, растворимость которого почти в 10 раз выше, чем мочевой кислоты. Действие препарата на фермент объясняется тем, что сначала он, подобно гипоксанти-ну, окисляется в гидроксипуринол, но при этом остаётся прочно связанным с активным центром фермента, вызывая его инактивацию;
с другой стороны, будучи псевдосубстратом, аллопуринол может превращаться в нуклеотид по "запасному" пути и ингибировать ФРДФ синтетазу и амидофосфорибозилтрансферазу, вызывая торможение синтеза пуринов de novo.
При лечении аллопуринолом детей с синдромом Лёша-Нихена удаётся предотвратить развитие патологических изменений в суставах и почках, вызванных гиперпродукцией мочевой кислоты, но препарат не излечивает аномалии в поведении, неврологические и психические расстройства.
Г. Гипоурикемия
Гипоурикемия и возросшая экскреция гипоксантина и ксантина может быть следствием недостаточности ксантиноксидазы, вызванной нарушениями в структуре гена этого фермента, либо результатом повреждения печени.
Биосинтез пиримидиновых рибонуклеотидов и его регуляция. Оротацидурия. «Запасные» пути синтеза пиримидиновых оснований.
V. БИОСИНТЕЗ ПИРИМИДИНОВЫХ НУКЛЕОТИДОВ
Фонд пирймидиновых нуклеотидов, подобно пуриновым нуклеотидам, в основном синтезируется из простых предшественников de novo, и только 10-20% от общего количества образуется по "запасным" путям из азотистых оснований или нуклеозидов.
А. Образование пиримидиновых нуклеотидов DE NOVO
В отличие от синтеза пуринов, где формирование гетероциклического основания осуществляется на остатке рибозо-5-фосфата, пиримидиновое кольцо синтезируется из простых предшественников: глутамина, СО2 и аспарагиновой кислоты и затем связывается с рибозо-5-фосфатом, полученным от ФРДФ.
Процесс протекает в цитозоле клеток. Синтез ключевого пиримидинового нуклеотида - УМФ идёт с участием 3 ферментов, 2 из которых полифункциональны.
Образование дигидрооротата
У млекопитающих ключевой, регуляторной реакцией в синтезе пирймидиновых нуклеотидов является синтез карбамоилфосфата из глутамина, СО2 и АТФ, в реакции катализируемой кар-бамоилфосфатсинтетазой II (КФС II), которая протекает в цитозоле клеток (рис. 10-12). В реакции NH2-гpyппa карбамоилфосфата образуется за счёт амидной группы глутамина, что отличает эту реакцию от реакции синтеза карбамоилфосфата в митохондриях в процессе синтеза мочевины из СО2, NH3 и АТФ с участием КФС I.
Карбамоилфосфат, использующийся на образование пирймидиновых нуклеотидов, является продуктом полифункционального фермента, который наряду с активностью КФС II содержит
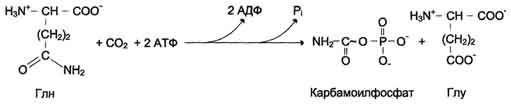
Рис. 10-12. Синтез карбамоилфосфата.
каталитические центры аспартаттранскарбамоилазы и дигидрооротазы. Этот фермент назвали "КАД-фермент" - по начальным буквам ферментативных активностей, которыми обладают отдельные каталитические домены этого белка. Объединение первых трёх ферментов метаболического пути в единый полифункциональный комплекс позволяет использовать почти весь синтезированный в первой реакции карбамоилфос-фат на взаимодействие с аспартатом и образование карбамоиласпартата, от которого отщепляется вода и образуется циклический продукт - дигидрооротат (рис. 10-13).
Отщепляясь от КАД-фермента, дигидрооротат подвергается дегидрированию NAD-зависимой дигидрооротатдегидрогеназой и превращается в свободное пиримидиновое основание - оротовую кислоту, или оротат.
Образование УМФ
В цитозоле оротат становится субстратом бифункционального фермента - УМФ-синтазы, которая обнаруживает оротатфосфорибозилтранс-феразную и ОМФ-декарбоксилазную активности. Первоначально фосфорибозильный остаток от ФРДФ переносится на оротат и образуется нук-леотид - оротидин-5'-монофосфат (ОМФ), декарбоксилирование которого даёт уридин-5-монофосфат (УМФ).
Таким образом, шесть последовательных реакций синтеза пиримидиновых нуклеотидов осуществляются тремя ферментами, которые кодируются в геноме человека тремя различными структурными генами.
Биосинтез УДФ, УТФ и иитидиловых нуклеотидов
УМФ под действием специфических нуклео-зидмонофосфат (НМФ) и нуклеозиддифосфат (НДФ) киназ превращается в УДФ и УТФ в результате переноса γ-фосфатного остатка АТФ на соответствующий субстрат.
НМФ-киназа катализирует следующую реакцию:
УМФ + АТФ → УДФ + АДФ,
а НДФ-киназа:
УДФ + АТФ → УТФ + АДФ.
ЦТФ синтетаза катализирует амидирование УТФ (рис, 10-14), осуществляя АТФ-зависимое замещение кетогругшы урацила на амидную группу глутамина с образованием цитидин-5'-трифосфата (ЦТФ).
Б. "Запасные" пути синтеза пиримидиновых нуклеотидов
Использование пиримидиновых оснований и нуклеозидов в реакциях реутилизации препятствует катаболизму этих соединений до конечных продуктов с расщеплением пиримидинового кольца. В ресинтезе пиримидинов участвуют некоторые ферменты катаболизма нуклеотидов. Так, уридинфосфорилаза в обратимой реакции может рибозилироватъ урацил с образованием уридина.
Урацил + Рибозо-1-фосфат → Уридин + Н3РО4.
Превращение нуклеозидов в нуклеотиды катализирует уридин-цитидинкиназа.
Часть ЦМФ может превращаться в УМФ под действием цитидиндезаминазы и пополнять запасы уридиловых нуклеотидов.
ЦМФ + Н2О → УМФ + NH3.
В. Регуляция синтеза пиримидиновых НУКЛЕОТИДОВ
Регуляторным ферментом в синтезе пиримидиновых нуклеотидов является полифункциональный

Рис. 10-13. Биосинтез УМФ de novo.

Рис. 10-14. Синтез ЦТФ из УТФ.
КАД-фермент. УМФ и пуриновые нуклеотиды аллостерически ингибируют, а ФРДФ активирует его карбамоилсинтетазную активность, тогда как активность аспартаттранскарбамоилазного домена ингибирует ЦТФ, но активирует АТФ (рис. 10-15).
Этот способ регуляции позволяет предотвратить избыточный синтез не только УМФ, но и всех других пиримидиновых нуклеотидов и обеспечить сбалансированное образование всех четырёх основных пуриновых и пиримидиновых нуклеотидов, необходимых для синтеза РНК.

Рис. 10-15. Регуляция синтеза пиримидиновых нуклеотидов. КАД-фермент катализирует реакции 1, 2, 3; дигидрооротатдегидрогеназа - реакцию 4; УМФ синтетаза - реакции 5 и 6; НМФ киназа - реакцию 7; НДФ киназа - реакцию 8; ЦТФ синтетаза - реацию 9.
VI. КАТАБОЛИЗМ ПИРИМИДИНОВЫХ НУКЛЕОТИДОВ
Уже говорилось о том, что цитидиловые нук-леотиды могут гидролитически терять аминогруппу и превращаться в УМФ. Когда от УМФ при участии нуклеотидазы (или фосфатазы) и уридинфосфорилазы отщепляются неорганический фосфат и рибоза, то остаётся азотистое основание - урацил. Аналогично расщепляются дезоксирибонуклеотиды, и из dЦМФ образуется урацил, а из dTМФ - тимин (рис. 10-16).
Пиримидиновые основания при участии дигидропиримидиндегидрогеназы присоединяют 2 атома водорода по двойной связи кольца с образованием дигидроурацила или дигидротимина. Оба гетероцикла могут взаимодействовать с
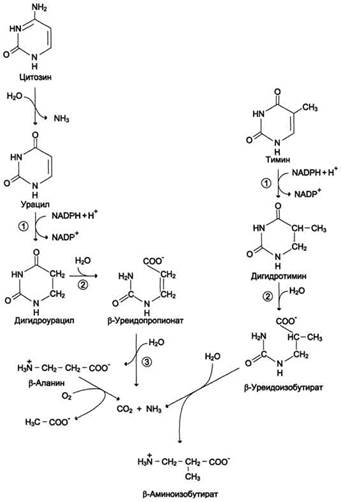
Рис. 10-16. Катаболизм пиримидиновых оснований. 1 - дигидропиримидиндегидрогеназа; 2 - дигидропиримидинциклогидролаза; 3 - уреидопропионаза.
водой в реакции, катализируемой дигидропиримидинциклогидролазой, и дигидроурацил превращается в β-уреидопропионовую кислоту, а дигидротимин - в β-уреидоизомасляную кислоту. Оба β-уреидопроизводных под действием общего для них фермента уреидопропионазы расщепляются с образованием СО2, NH4+ и β-аланина или β-аминоизомасляной кислоты соответственно.
β-Аланин обнаруживают в плазме крови и многих тканях. Он используется в мышцах на образование дипептидов: карнозина и анзерина. Под действием бактериальной микрофлоры кишечника β-аланин включается в пантотеновую кислоту, которая всасывается и используется на образование КоА.
Часть β-аланина и β-аминбутирата трансами-нируется с α-кетоглутаратом и даёт малонил полуальдегид или метилмалонил полуальдегид соответственно, которые превращаются в малонил-КоА и сукцинил-КоА и используются в соответствующих метаболических путях, либо окисляются до СО2 и Н2О. Частично β-аминобутират экскретируется с мочой.
VII. НАРУШЕНИЯ ОБМЕНА ПИРИМИДИНОВЫХ НУКЛЕОТИДОВ
Описано несколько нарушений, связанных со снижением активности ферментов обмена пиримидиновых нуклеотидов. Одно из них - оротацидурия - вызвано дефектом в работе второго бифункционального фермента синтеза нуклеотидов de novo - УМФ-синтазы, два других обнаружены в процессе катаболизма пиримидинов.
А. Оротацидурия
Это единственное нарушение синтеза пиримидинов de novo. Оно вызвано снижением активности УМФ-синтазы, которая катализирует образование и декарбоксилирование ОМФ. Поскольку в эмбриогенезе от образования пиримидинов de novo зависит обеспечение синтеза ДНК субстратами, то жизнь плода невозможна при полном отсутствии активности этого фермента. Действительно, у всех пациентов с оротацидурией отмечают заметную, хотя и очень низкую активность УМФ-синтазы. Установлено, что содержание оротовои кислоты в моче пациентов (1 г/сут и более) значительно превосходит количество оротата, которое ежедневно синтезируется в норме (около 600 мг/сут). Снижение синтеза пиримидиновых нуклеотидов, наблюдающееся при этой патологии, нарушает регуляцию КАД-фермента по механизму ретроингибирования, из-за чего возникает гиперпродукция оротата.
Клинически наиболее характерное следствие оротацидурии - мегалобластная анемия, вызванная неспособностью организма обеспечить нормальную скорость деления клеток эритроцитарного ряда. Её диагностируют у детей на том основании, что она не поддаётся лечению препаратами фолиевой кислоты.
Недостаточность синтеза пиримидиновых нуклеотидов сказывается на интеллектуальном развитии, двигательной способности и сопровождается нарушениями работы сердца и ЖКТ. Нарушается формирование иммунной системы, и наблюдается повышенная чувствительность к различным инфекциям.
Гиперэкскреция оротовои кислоты сопровождается нарушениями со стороны мочевыводя-щей системы и образованием камней. При отсутствии лечения больные обычно погибают в первые годы жизни. При этом оротовая кислота не оказывает токсического эффекта. Многочисленные нарушения в работе разных систем организма вызваны "пиримидиновым голодом".
Для лечения этой болезни применяют уридин (от 0,5 до 1 г/сут), который по "запасному" пути превращается в УМФ.
Уридин + АТФ → УМФ + АДФ.
Нагрузка уридином устраняет "пиримидиновый голод", а поскольку из УМФ могут синтезироваться все остальные нуклеотиды пиримидинового ряда, то снижается выделение оротовои кислоты из-за восстановления механизма ретроингибирования КАД-фермента. Для больных оротацидурией лечение уридином продолжается в течение всей жизни, и этот нуклеозид становится для них незаменимым пищевым фактором.
Кроме генетически обусловленных причин, оротацидурия может наблюдаться:
при гипераммониемии, вызванной дефектом любого из ферментов орнитинового цикла,
за исключением карбамоилфосфат- синтетазы I. В этом случае карбамоилфосфат, синтезированный в митохондриях, выходит в цитозоль клеток и начинает использоваться на образование пиримидиновых нуклеотидов. Концентрация всех метаболитов, в том числе и оротовой кислоты, повышается. Наиболее значительная экскреция оротата отмечается при недостаточности орнитинкарбамоилтрансферазы (второго фермента орнитинового цикла);
в процессе лечения подагры аллопуринолом, который превращается в оксипуринолмононуклеотид и становится сильным ингибитором УМФ-синтазы. Это приводит к накоплению оротовой кислоты в тканях и крови.
Б. Нарушения катаболизма пиримидинов
Известны нарушения в работе 2 ферментов этого метаболического пути.
При недостаточности пиримидин-5'-нуклеотидазы нарушаются отщепление неорганического фосфата от пиримидиновых мононуклеотидов и образование нуклеозидов.
Неактивная изоформа пиримидин-5'-нуклеотидазы обнаружена в эритроцитах. В результате наблюдается накопление гиримидиновых НТФ, которые ингибируют пентозофосфатный путь превращения глюкозы и тем самым создают предпосылки к гемолизу эритроцитов (см. раздел 14).
Дигидропиримидиндегидрогеназа - скорость-лимитирующий фермент катаболизма пиримидинов. Нарушение работы этого фермента сопровождается отклонениями в функционировании нервной системы и диагностируется на основании повышения уровня свободных пиримидинов: урацила и тимина в плазме крови.
Биосинтез дезоксирибонуклеотидов с помощью рибонуклеотидредуктазного комплекса. Образование дТМФ, роль Н4-фолата в этом процессе. Ингибиторы синтеза дезоксирибонуклеотидов как лекарственные препараты: 5- фторурацил, азитотимидин и др.
VIII. БИОСИНТЕЗ ДЕЗОКСИРИБОНУКЛЕОТИДОВ
Синтез дезоксирибонуклеотидов идёт с заметной скоростью только в тех клетках, которые вступают в S-фазу клеточного цикла и готовятся к синтезу ДНК и делению. В покоящихся клетках дезоксинуклеотиды практически отсутствуют. Все дезоксинуклеотиды, кроме тимидиловых, образуются из рибонуклеотидов путём прямого восстановления ОН-группы у второго углеродного атома рибозы в составе рибонуклеозиддифосфатов до дезоксирибозы. Тимидиловые нук-леотиды синтезируются из dУМФ особым путём с участием N5,N10-метилен-Н4-фолата.
А. Рибонуклеотидредуктазный комплекс
Реакцию восстановления НДФ в дезоксипроизводные катализирует рибонуклеотидредук-тазный комплекс, в состав которого входят: собственно рибонуклеотидредуктаза (РНР), белок тиоредоксин и фермент тиоредоксинредуктаза, обеспечивающий регенерацию восстановленной формы тиоредоксина (рис. 10-17).
Рибонуклеотидредуктаза - олигомерный белок, состоящий из двух В1- и двух В2-субъединиц, и содержит негеминовое железо в качестве кофактора.
Непосредственным донором водорода в реакции восстановления рибозы служит низкомолекулярный белок тиоредоксин. В рабочую часть этого белка входят 2 SH-группы, которые, отдавая водород, окисляются с образованием дисульфидного мостика. Второй фермент комплекса - тиоредоксинредуктаза - катализирует гидрирование окисленного тиоредоксина с использованием NADPH.
При участии комплекса РНР образуются: dАДФ, dГДФ, dУДФ и dЦДФ, которые с помощью НДФ-киназ превращаются в дНТФ, 3 из которых (кроме дУДФ) непосредственно используются в синтезе ДНК.
дНДФ + АТФ → дНТФ + АДФ.
Б. Биосинтез тимидиловых нуклеотидов
Тимидин-5'-монофосфат (дТМФ) образуется из дУМФ в реакции, катализируемой тимиди-латсинтазой (рис. 10-18). Донором метильной группы, появляющейся в 5-положении пиримидинового кольца в молекуле дТМФ, служит кофермент тимидилатсинтазы - N5,N10-метилен-Н4-фолат. С помощью этого кофермента в молекулу дУМФ включается метиленовая группа и восстанавливается в метальную, используя 2 атома водорода от Н4-фолата.

Рис. 10-17. Восстановление рибонуклеозиддифосфатов в 2'-дезоксирибонуклеозиддифосфаты. 1 - рибонуклеотидредуктаза (РНР); 2 - тиоредоксинредуктаза.
Образование субстрата тимидилатсинтазной реакции - дУМФ осуществляется двумя путями (рис. 10-19):
дефосфорилированием дУДФ;
гидролитическим дезаминированием дЦМФ с помощью дЦМФ дезаминазы. дЦМФ получается при дефосфорилировании дЦДФ - одного из продуктов рибонуклеотидредуктаз-ной реакции. В организме человека это основной путь образования дУМФ.
Скорость синтеза дТМФ зависит также от количества второго субстрата тимидилатсинтазной реакции - N5,N10-метилен-Н4-фолата, пополнение запасов которого осуществляется при участии 2 ферментов: дигидрофолатредуктазы, которая с участием NADPH восстанавливает Н2-фолат в Н4-фолат, и серии гидроксиметилтрансферазы, осуществляющей перенос β-гидроксиметиленовой группы серина на Н4-фолат (см. раздел 9). У человека дТМФ образуется, главным образом, из дЦДФ.
В. "Запасные" пути синтеза дезоксирибонуклеотидов
В быстроделящихся клетках наряду с синтезом дезоксинуклеотидов с помощью рибонуклеотид-редуктазного комплекса и тимидилатсинтазы активируются реакции, обеспечивающие повторное использование тимина и дезоксицитидина в реакциях, катализируемых ферментами "запасных" путей и обратимых реакций катаболизма. Под влиянием тимидинфосфорилазы протекает следующая реакция:
Тимин + Дезоксирибоза-1-фосфат → Тимидин + Н3Р04.

Рис. 10-19. Образование ТТФ из дЦДФ и дУДФ. 1 - дЦМФ дезаминаза; 2 -тимидилатсинтаза; 3 - дНМ
|
|
|
