 |


Глава 8. Соединительные ткани
|
|
|
|
Соединительные ткани - это большая группа тканей производных мезенхимы, как правило, полидифферонные и с преобладанием межклеточного вещества (волокнистых структур и аморфного компонента), участвующие в поддержании постоянства состава внутренней среды организма.
Соединительная ткань составляет более 50 % массы тела человека. Она участвует в формировании стромы органов, прослоек между другими тканями, дермы кожи, скелета. Полифункциональный характер соединительных тканей определяется сложностью их состава и организации.
Функции соединительных тканей. Соединительные ткани выполняют различные функции: трофическую, защитную, опорную (биомеханическую), пластическую, морфогенетическую. Трофическая функция в широком смысле этого слова связана с регуляцией питания различных тканевых структур, с участием в обмене веществ и поддержанием постоянства состава внутренней среды организма. В реализации этой функции главную роль играет основное вещество, через которое осуществляется транспорт воды, солей, молекул питательных веществ, - интегративно-буферная среда. Защитная функция заключается в предохранении организма от нефизиологических механических воздействий (повреждений) и обезвреживании чужеродных веществ, поступающих извне или образующихся внутри организма. Это обеспечивается физической защитой (костной тканью), а также фагоцитарной деятельностью макрофагов и иммунокомпе-тентными клетками, участвующими в реакциях клеточного и гуморального иммунитета. Опорная (биомеханическая) функция обеспечивается прежде всего коллагеновыми и эластическими волокнами, образующими волокнистые основы всех органов, составом и физико-химическими свойствами межклеточного вещества скелетных тканей (минерализацией). Чем плотнее межклеточное вещество, тем значительнее опорная, биомеханическая функция. Пластическая функция соединительной ткани выражается в адаптации к меняющимся условиям существования, регенерации, участии в замещении дефектов органов при их повреждении. Морфогенетическая
|
|
|
(структурообразовательная) функция проявляется формированием тканевых комплексов и обеспечением общей структурной организации органов (образование капсул, внутриорганных перегородок), регулирующим влиянием некоторых ее компонентов на пролиферацию и дифференцировку клеток различных тканей.
Классификация соединительных тканей. Разновидности соединительной ткани различаются по составу и соотношению клеточных дифферонов, волокон, а также по физико-химическим свойствам аморфного межклеточного вещества. Соединительные ткани подразделяются на собственно соединительную ткань (рыхлая соединительная ткань и соединительные ткани со специальными свойствами) и скелетные ткани. Последние, в свою очередь, подразделяются на три разновидности хрящевой ткани (гиалиновая, эластическая, волокнистая), две разновидности костной ткани (ретикулофиброзная и пластинчатая), а также цемент и дентин зуба (схема 8.1).
Гистогенез соединительных тканей. Источником развития соединительных тканей является мезенхима (от греч. mesos - средний, enchima - заполняющая масса). Это один из эмбриональных зачатков (по некоторым представлениям - эмбриональная ткань), представляющий собой разрыхленную честь среднего зародышевого листка. Клеточные элементы мезенхимы образуются в процессе дифференцировки дерматома, склеро-тома, висцерального и париетального листков спланхнотома. Кроме того, существует эктомезенхима (нейромезенхима), развивающаяся из нервного гребня (ганглиозной пластинки). По мере развития зародыша в мезенхиму мигрируют клетки иного происхождения из других эмбриональных
|
|
|
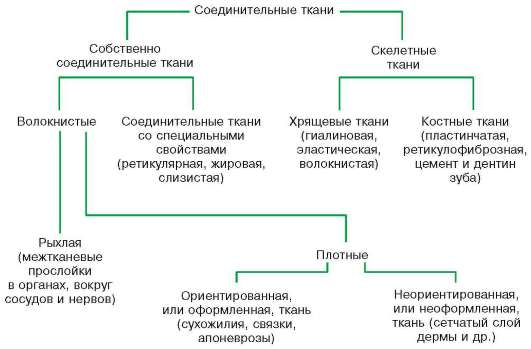
Схема 8.1. Классификация соединительных тканей
зачатков, например клетки нейробластического дифферона, миобласты из закладки скелетных мышц, пигментоциты и др.
Следовательно, с определенной стадии развития зародыша мезенхима представляет собой мозаику клеток, возникших из разных зародышевых листков и эмбриональных зачатков тканей. Однако морфологически все клетки мезенхимы мало чем отличаются друг от друга, и только очень чувствительные методы исследования (как правило, иммуноцитохимические, электронно-микроскопические) выявляют в составе мезенхимы клетки различной природы. Мезенхима существует только в эмбриональном периоде развития человека. После рождения в организме человека сохраняются лишь малодифференцированные (полипотентные) клетки в составе рыхлой волокнистой соединительной ткани (адвентициальные клетки), которые могут дивергентно дифференцироваться в различных направлениях, но в пределах определенной тканевой системы.
Различают эмбриональный и постэмбриональный гистогенез соединительных тканей. В процессе эмбрионального гистогенеза мезенхима приобретает черты тканевого строения раньше закладки других тканей. Этот процесс в различных органах и системах происходит неодинаково и зависит от их неодинаковой физиологической значимости на различных этапах эмбриогенеза. В дифференцировке мезенхимы отмечаются топографическая асин-хронность как в зародыше, так и во внезародышевых органах, высокие темпы размножения клеток и волокнообразования. Постэмбриональный гистогенез в нормальных физиологических условиях происходит медленнее и направлен на поддержание тканевого гомеостаза, пролиферацию малодифференциро-ванных клеток и замену ими отмирающих клеток. Существенную роль в этих процессах играют межклеточные внутритканевые взаимодействия, индуцирующие и ингибирующие факторы (интегрины, межклеточные адгезивные факторы, функциональные нагрузки, гормоны, оксигенация, наличие мало-дифференцированных клеток).
Общие принципы организации соединительных тканей. Главными компонентами соединительных тканей являются производные клеток - волокнистые структуры коллагенового и эластического типов, основное (аморфное) вещество, играющее роль интегративно-буферной метаболической среды, и клеточные элементы, создающие и поддерживающие количественное и качественное соотношение состава неклеточных компонентов.
|
|
|
Органная специфичность клеточных элементов соединительной ткани выражается в количестве, форме и соотношении различных видов клеток, их метаболизме и функциях, оптимально приспособленных к функции органа. Специфичность клеточных элементов проявляется также их взаимодействием между собой (индивидуально расположенные, клеточные ассоциации), особенностями их внутреннего строения (состав органелл, структура ядра, наличие ферментов и др.). Специфика соединительной ткани обнаруживается и в соотношении клеток различных дифферонов и неклеточных структур в различных участках тела. В рыхлой волокнистой соединительной ткани превалируют клетки различных дифферонов и аморфное вещество над волокнами, а в плотной соединительной ткани - наоборот, основную ее массу составляют волокна.
8.1. СОБСТВЕННО СОЕДИНИТЕЛЬНАЯ ТКАНЬ
8.1.1. Волокнистые соединительные ткани
Рыхлая соединительная ткань
Рыхлая соединительная ткань (textus connectivus laxus) обнаруживается во всех органах, так как она сопровождает кровеносные и лимфатические сосуды и образует строму многих органов. Несмотря на наличие органных особенностей, строение рыхлой соединительной ткани в различных органах имеет сходство. Она состоит из клеток различной гистогенетической детерминации и межклеточного вещества (рис. 8.1).

Рис. 8.1. Рыхлая соединительная ткань:
а - пленочный препарат: 1 - фибробласт; 2 - макрофаги; 3 - коллагеновые волокна; 4 - эластические волокна; 5 - лимфоцит; б - фибробласт; в - макрофаг

Рис. 8.2. Ультрамикроскопическое строение фибробласта на разных стадиях дифференцировки (по Н. А. Юриной и А. И. Радостиной, с изменениями): Фибробласты: а - малодифференцированный; б - молодой; в - зрелый; г - фиброцит. 1 - ядро; 2 - комплекс Гольджи; 3 - митохондрии; 4 - рибосомы и полирибосомы; 5 - гранулярная эндоплазматическая сеть; 6 - коллагеновые фибриллы
|
|
|
Клетки
Основными клетками соединительной ткани являются фибробласты (семейство фибриллообразующих клеток), макрофаги (семейство), тучные клетки, адвентициальные клетки, плазматические клетки, перициты, жировые клетки, а также лейкоциты, мигрирующие из крови; иногда пигментные
клетки.
Фибробласты (от лат. fibra - волокно, греч. blastos - росток, зачаток) - клетки, синтезирующие компоненты межклеточного вещества: белки (коллаген, эластин), протеогликаны, гликопротеины (см. рис. 8.1; рис. 8.2, 8.3).
Среди мезенхимных клеток имеются стволовые клетки, дающие начало дифферону фибробластов: стволовые клетки, полустволовые клетки-предшественники, малодифференцированные (малоспециализированные), дифференцированные фибробласты (зрелые, активно функционирующие), фиброциты (дефинитивные - конечные формы клеток), а также миофиб-робласты и фиброкласты. С главной функцией фибробластов связаны образование основного вещества и волокон, заживление ран, развитие грануляционной ткани, образование соединительнотканной капсулы вокруг инородного тела и др. Морфологически в этом диффероне можно идентифицировать только клетки, начиная с малодифференцированного фибробласта. Последние - малоотростчатые клетки с округлым или овальным ядром и небольшим ядрышком, базофильной цитоплазмой, богатой РНК. Размер клеток не превышает 20-25 мкм. В цитоплазме этих клеток обнаруживается
большое количество свободных рибосом (см. рис. 8.2). Эндоплазматическая сеть и митохондрии развиты слабо. Комплекс Гольджи представлен скоплениями коротких трубочек и пузырьков.
Авторадиографически показано, что на этой стадии цитогенеза фибробласты обладают очень низким уровнем синтеза и секреции белка. Эти фибробласты способны к размножению митотическим путем.
Дифференцированные зрелые фибробласты крупнее по размеру и в распластанном виде на пленочных препаратах могут достигать 40-50 мкм и более (см. рис. 8.1). Это активно функционирующие клетки. Ядра у них светлые, овальные, содержат 1-2 крупных ядрышка; цитоплазма базофильна, с хорошо развитой гранулярной эндоплазматической сетью, которая местами контактирует с плазмолеммой (см. рис. 8.2, рис. 8.3, а). Комплекс Гольджи распределен в виде цистерн и пузырьков по всей клетке. Митохондрии и лизосомы развиты умеренно.
В цитоплазме фибробластов, особенно в периферической части, располагаются микрофиламенты толщиной 5-6 нм, содержащие белки типа актина и миозина, что обусловливает способность этих клеток к движению. Движение фибробластов становится возможным только после их связывания с опорными фибриллярными структурами (фибрин, соединительнотканные волокна) с помощью фибронектина - гликопротеина, синтезированного фибробластами и другими клетками, обеспечивающего адгезию клеток и неклеточных структур. Во время движения фибробласт уплощается, а его поверхность может увеличиться в 10 раз.
|
|
|
Плазмолемма фибробластов является важной рецепторной зоной, которая опосредует воздействие различных регуляторных факторов. Активизация фибробластов обычно сопровождается накоплением гликогена и повышенной активностью гидролитических ферментов. Энергия, образуемая при метаболизме гликогена, используется для синтеза полипептидов и других компонентов, секретируемых клеткой.
Биосинтез белков коллагена и эластина, протеогликанов, необходимых для формирования основного вещества и волокон, в зрелых фибробластах осуществляется довольно интенсивно, особенно в условиях пониженной концентрации кислорода. Стимулирующими факторами биосинтеза коллагена являются ионы железа, меди, хрома, аскорбиновая кислота. Один из гидролитических ферментов - коллагена-за - расщепляет внутри клеток незрелый коллаген, что, по-видимому, регулирует на клеточном уровне интенсивность секреции коллагена.
По способности синтезировать фибриллярные белки к семейству фибробластов можно отнести ретикулярные клетки ретикулярной соединительной ткани кроветворных органов, хондробласты и остеобласты скелетных разновидностей соединительной ткани.
Фиброциты - дефинитивные формы развития фибробластов. Эти клетки веретенообразные с крыловидными отростками. Они содержат небольшое число органелл, вакуолей, липидов и гликогена. Синтез коллагена и других веществ в фиброцитах резко снижен.
Миофибробласты - клетки, морфологически сходные с фибробластами, сочетающие в себе способность к синтезу не только коллагена, но и сократи-

Рис. 8.3. Фибробласт, миофибробласт и фиброкласт:
а - фибробласт (препарат А. И. Радостиной): 1 - ядро; 2 - гранулярная эндоплаз-матическая сеть; 3 - рибосомы; 4 - комплекс Гольджи; 5 - митохондрии; 6 - плаз-молемма; 7 - коллагеновые фибриллы; б - миофибробласт (препарат А. Б. Шехтера): 1 - ядро; 2 - гранулярная эндоплазматическая сеть; 3 - рибосомы; 4 - комплекс Гольджи; 5 - сократительные филаменты; 6 - плазмолемма
тельных белков в значительном количестве (см. рис. 8.3, б). Установлено, что фибробласты могут развиваться в миофибробласты, функционально сходные с гладкими мышечными клетками, но в отличие от последних имеют хорошо развитую эндоплазматическую сеть. Такие клетки наблюдаются в
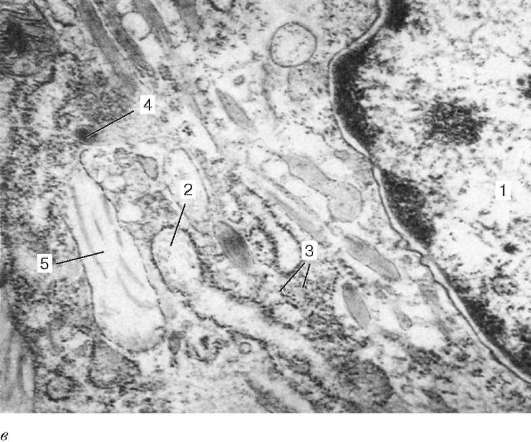
Рис. 8.3. Продолжение:
в - фиброкласт (препарат А. Б. Шехтера): 1 - ядро; 2 - гранулярная эндоплазмати-ческая сеть; 3 - рибосомы; 4 - лизосомы; 5 - фаголизосомы с фрагментами колла-геновых фибрилл. Электронные микрофотографии. Увеличение 20 000
грануляционной ткани в условиях регенерации раневого процесса и в матке при развитии беременности.
Фиброкласты - клетки с высокой фагоцитарной и гидролитической активностью, принимают участие в «рассасывании» межклеточного вещества (см. рис. 8.3, в) в период инволюции органов (например, матки после окончания беременности). Они сочетают в себе структурные признаки фибриллообразующих клеток (развитую гранулярную эндоплазматическую сеть, комплекс Гольджи, относительно крупные, но немногочисленные митохондрии), а также лизосомы с характерными для них гидролитическими ферментами. Выделяемый ими за пределы клетки комплекс ферментов расщепляет цементирующую субстанцию коллагеновых волокон, после чего происходят фагоцитоз и внутриклеточное переваривание коллагена кислыми протеазами лизосом.
Макрофаги (макрофагоциты) (от греч. makros - большой, длинный, fagos - пожирающий) - это гетерогенная специализированная клеточная популяция защитной системы организма. Различают две группы макрофагов - свободные и фиксированные. К свободным макрофагам относятся макрофаги рыхлой соединительной ткани, или гистиоциты; макрофаги серозных полостей; макрофаги воспалительных экссудатов; альвеолярные макрофаги легких. Макрофаги способны перемещаться в организме. Группу фиксированных (резидентных) макрофагов составляют макрофаги костного мозга, костной и хрящевой тканей (остеокласты, хондрокласты), селе-
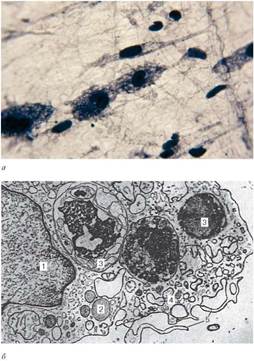
Рис. 8.4. Макрофаги:
а - макрофаги подкожной соединительной ткани крысы (микрофотография, окраска - железный гематоксилин); б - макрофаг (электронная микрофотография; препарат А. И. Радостиной, увеличение 18 000): 1 - ядро; 2 - первичные лизосомы; 3 - вторичные лизосомы; 4 - профили канальцев эндоплазматической сети; 5 - микровыросты периферического слоя цитоплазмы
зенки, лимфатических узлов (дендритные макрофаги), внутриэпидермаль-ные макрофаги (клетки Лангерганса), макрофаги ворсин плаценты (клетки Хофбауэра), макрофаги ЦНС (микроглия).
Размер и форма макрофагов варьируют в зависимости от их функционального состояния (рис. 8.4). Обычно макрофаги, за исключением некоторых их
видов (гигантские клетки инородных тел, хондро- и остеокласты), имеют одно ядро. Ядра макрофагов небольшого размера, округлые, бобовидные или неправильной формы. В них содержатся крупные глыбки хроматина. Цитоплазма базофильна, богата лизосомами, фагосомами (отличительные признаки) и пиноцитозными пузырьками, содержит умеренное количество митохондрий, гранулярную эндоплазматическую сеть, комплекс Гольджи, включения гликогена, липидов и другие (см. рис. 8.4, б). В цитоплазме макрофагов выделяют «клеточную периферию», обеспечивающую макрофагу способность передвигаться, втягивать микровыросты цитоплазмы, осуществлять эндо- и экзоци-тоз. Непосредственно под плазмолеммой находится сеть актиновых филамен-тов диаметром 5-6 нм. Через эту сеть проходят микротрубочки диаметром 20 нм, которые прикрепляются к плазмолемме. Микротрубочки направлены радиально от клеточного центра к периферии клетки и играют важную роль во внутриклеточных перемещениях лизосом, микропиноцитозных везикул и других структур. На поверхности плазмолеммы имеются рецепторы для опухолевых клеток и эритроцитов, Т- и В-лимфоцитов, антигенов, иммуноглобулинов, гормонов. Наличие рецепторов иммуноглобулинов обусловливает участие макрофагов в иммунных реакциях (см. главу 14).
Формы проявления защитной функции макрофагов: 1) поглощение и дальнейшее расщепление или изоляция чужеродного материала; 2) обезвреживание его при непосредственном контакте; 3) передача информации о чужеродном материале иммунокомпетентным клеткам, способным его нейтрализовать; 4) оказание стимулирующего воздействия на другую клеточную популяцию защитной системы организма. Макрофаги имеют орга-неллы, синтезирующие ферменты для внутриклеточного и внеклеточного расщепления чужеродного материала, антибактериальные и другие биологически активные вещества (протеазы, кислые гидролазы, пироген, интерферон, лизоцим и др.).
Количество макрофагов и их активность особенно возрастают при воспалительных процессах. Макрофаги вырабатывают хемотаксические факторы для лейкоцитов. Секретируемый макрофагами ИЛ-1 способен повышать адгезию лейкоцитов к эндотелию, секрецию лизосомных ферментов ней-трофилами и их цитотоксичность, активирует синтез ДНК в лимфоцитах. Макрофаги вырабатывают факторы, активирующие выработку иммуноглобулинов В-лимфоцитами, дифференцировку Т- и В-лимфоцитов; цитоли-тические противоопухолевые факторы, а также факторы роста, влияющие на размножение и дифференцировку клеток собственной популяции, стимулируют функцию фибробластов (см. главу 14).
Контакт макрофагов с антигенами резко усиливает расход глюкозы, липидный обмен и фагоцитарную активность.
Макрофаги образуются из СКК, а также из промоноцита и моноцита (см. рис. 7.15). Полное обновление макрофагов рыхлой волокнистой соединительной ткани у экспериментальных животных осуществляется примерно в 10 раз быстрее, чем фибробластов.
Одной из разновидностей макрофагов являются многоядерные гигантские клетки, которые раньше называли «гигантскими клетками инород-
ных тел», так как они могут формироваться, в частности, в присутствии инородного тела. Многоядерные гигантские клетки содержат 10-20 ядер и более, возникают либо путем слияния одноядерных макрофагов, либо путем эндомитоза без цитотомии. По данным электронной микроскопии, в многоядерных гигантских клетках присутствуют синтетический и секреторный аппарат и большое количество лизосом. Плазмолемма образует многочисленные складки.
Понятие о макрофагической системе. К этой системе относятся макрофаги (гистиоциты) рыхлой волокнистой соединительной ткани, звездчатые клетки синусоидных сосудов печени, свободные и фиксированные макрофаги кроветворных органов (костного мозга, селезенки, лимфатических узлов), макрофаги легкого, воспалительных экссудатов (перитонеальные макрофаги), остеокласты, гигантские клетки инородных тел и глиальные макрофаги нервной ткани (микроглия). Все они активные фагоциты. Фагоцитированный материал подвергается внутри клетки ферментативному расщеплению («завершенный фагоцитоз»), благодаря чему ликвидируются вредные для организма агенты, возникающие местно или проникающие извне. Клетки имеют на своей поверхности рецепторы иммуноглобулинов и возникают из промоноци-тов костного мозга и моноцитов крови. В отличие от таких «профессиональных» фагоцитов способностью к факультативному поглощению обладают фибробласты, ретикулярные клетки, эндотелиоциты, нейтро-фильные гранулоциты и др. Но эти клетки не входят в состав макро-фагической системы.
И. И. Мечников первым пришел к мысли о том, что фагоцитоз, возникающий в процессе эволюции как форма внутриклеточного пищеварения и закрепившийся за многими клетками, одновременно является важным защитным механизмом. Он обосновал целесообразность объединения их в одну систему и предложил назвать ее макрофагиче-ской. Макрофагическая система представляет собой мощный защитный аппарат, принимающий участие как в общих, так и в местных защитных реакциях организма. В целостном организме макрофагиче-ская система регулируется как местными механизмами, так и нервной и эндокринной системами.
В 1930-1940-х гг. эту защитную систему называли ретикулоэндоте-лиальной. В последнее время ее называют системой мононуклеарных фагоцитов, что, однако, неточно характеризует ее в связи с тем, что среди клеток, входящих в эту систему, есть и многоядерные (остеокласты).
Тучные клетки (мастоциты, тканевые базофилы, лаброциты). Этими терминами называют клетки, в цитоплазме которых находится специфическая зернистость, напоминающая гранулы базофильных гранулоцитов. Тучные клетки принимают участие в понижении свертывания крови, повышении проницаемости гематотканевого барьера, в процессе воспаления, иммуно-
генеза и др. У человека тучные клетки обнаруживаются всюду, где имеются прослойки рыхлой соединительной ткани. Особенно много тучных клеток в стенке органов пищеварительного тракта, матке, молочной железе, тимусе, миндалинах. Они часто располагаются группами по ходу кровеносных сосудов микроциркуляторного русла - капилляров, артериол, венул и мелких лимфатических сосудов (рис. 8.5, а).
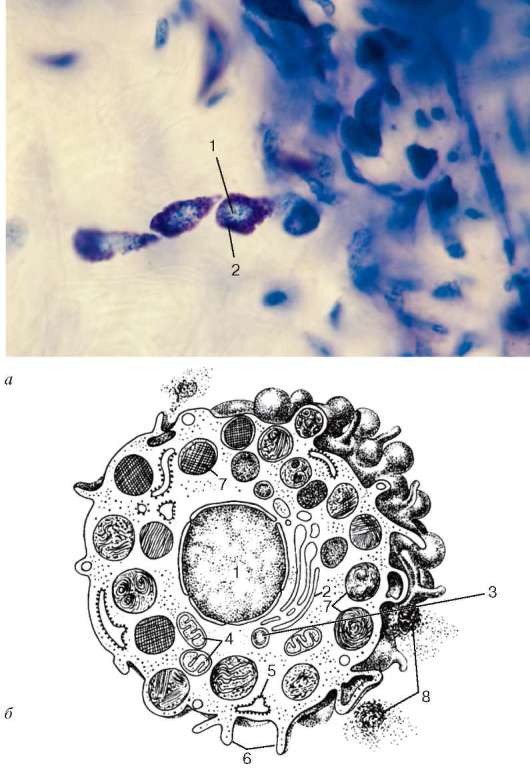
Рис. 8.5. Тучные клетки: а - в подкожной соединительной ткани (микрофотография): 1 - ядро; 2 - мета-хроматические гранулы в цитоплазме; б - схема ультрамикроскопического строения (по Ю. И. Афанасьеву): 1 - ядро; 2 - комплекс Гольджи; 3 - лизосома; 4 - митохондрии; 5 - эндоплазматическая сеть; 6 - микроворсинки; 7 - гетерогенные гранулы; 8 - секреторные гранулы в межклеточном веществе
Форма тучных клеток разнообразна. Клетки могут быть неправильной формы, овальными. Иногда эти клетки имеют короткие широкие отростки, что обусловлено способностью их к амебоидным движениям. У человека ширина таких клеток колеблется от 4 до 14 мкм, длина до 22 мкм. Ядра клеток сравнительно невелики, обычно круглой или овальной формы с плотно расположенным хроматином. В цитоплазме имеются многочисленные гранулы. Величина, состав и количество гранул варьируют. Их диаметр около 0,3-1 мкм (рис. 8.5, б). Меньшая часть гранул представляет собой ортохроматически окрашивающиеся азурофильные лизосомы. Большинство гранул отличаются метахромазией, содержат гепарин, хондроитинсерные кислоты типа А и С, гиалуроновую кислоту, гистамин, серотонин. Гранулы имеют сетчатое, пластинчатое, кристаллоидное и смешанное строение.
Органеллы тучных клеток (митохондрии, комплекс Гольджи, эндоплаз-матическая сеть) развиты слабо. В цитоплазме обнаружены различные ферменты: протеазы, липазы, кислая и щелочная фосфатазы, пероксидаза, цитохромоксидаза, АТФ-аза и др. Однако маркерным ферментом следует считать гистидиндекарбоксилазу, с помощью которой осуществляется синтез гистамина из гистидина.
Тучные клетки способны к секреции и выбросу своих гранул. Дегрануляция тучных клеток может происходить в ответ на любое изменение физиологических условий и действие патогенов. Однако выход биогенных аминов из тучной клетки может происходить и путем секреции растворимых компонентов (секреция гистамина). Гистамин немедленно вызывает расширение кровеносных капилляров и повышает их проницаемость, что проявляется локальными отеками. Он обладает также гипотензивным свойством и является важным медиатором воспаления.
Гистамин выступает как антагонист гепарина, который снижает проницаемость межклеточного вещества и свертываемость крови, оказывает противовоспалительное влияние.
Количество тучных клеток изменяется в зависимости от физиологических состояний организма: возрастает в матке, молочных железах в период беременности, а в желудке, кишечнике, печени - на высоте пищеварения.
Предшественники тучных клеток происходят из стволовых кроветворных клеток красного костного мозга. Процессы митотического деления тучных клеток наблюдаются крайне редко.
Плазматические клетки (плазмоциты). Эти клетки обеспечивают выработку антител - гамма-глобулинов (белки) при появлении в организме антигена. Они образуются в лимфоидных органах из В-лимфоцитов (см. главу 14), обычно встречаются в рыхлой соединительной ткани собственной пластинки слизистых оболочек полых органов, сальнике, интерстициальной соединительной ткани различных желез (молочных, слюнных и др.), лимфатических узлах, селезенке, костном мозге и др.
Величина плазмоцитов колеблется от 7 до 10 мкм. Форма клеток круглая или овальная. Ядра относительно небольшие, круглой или овальной формы, расположены эксцентрично. Цитоплазма резко базофильна, содержит хорошо развитую концентрически расположенную гранулярную эндоплазма-

Рис. 8.6. Адипоциты:
1 - капилляр; 2 - липидные включения; 3 - ядро; 4 - цитоплазма (микрофотография). Окраска - железный гематоксилин
тическую сеть, в которой синтезируются белки (антитела). Базофилия отсутствует только в небольшой светлой зоне цитоплазмы около ядра, образующей так называемую сферу или дворик. Здесь располагаются центриоли и комплекс Гольджи. Иногда обнаруживаются скопления иммуноглобулинов в виде оксифильных телец Русселя.
Для плазматических клеток характерна высокая скорость синтеза и секреции антител, что отличает их от своих предшественников. Хорошо развитый секреторный аппарат позволяет синтезировать и секретировать несколько тысяч молекул иммуноглобулинов в секунду. Количество плаз-моцитов увеличивается при различных инфекционно-аллергических и воспалительных заболеваниях.
Адипоциты (жировые клетки). Так называют клетки, которые обладают способностью накапливать в больших количествах резервный жир, принимающий участие в трофике, энергообразовании и метаболизме воды. Адипоциты располагаются группами, реже поодиночке и, как правило, около кровеносных сосудов. Накапливаясь в больших количествах, эти клетки образуют жировую ткань (белую или бурую) (см. ниже).
Форма одиночно расположенных жировых клеток шаровидная. Зрелая жировая клетка обычно содержит одну большую каплю нейтрального жира (триглицеридов), занимающую всю центральную часть клетки и окруженную тонким цитоплазматическим ободком, в утолщенной части которого лежит ядро (рис. 8.6). Кроме того, в цитоплазме адипоцитов имеется небольшое количество других липидов: холестерина, фосфолипидов, свободных жирных кислот и др. Липиды хорошо окрашиваются суданом III в оранжевый цвет или осмиевой кислотой в черный цвет. В прилежащей к ядру цитоплазме, а иногда и в более тонкой противоположной ее части
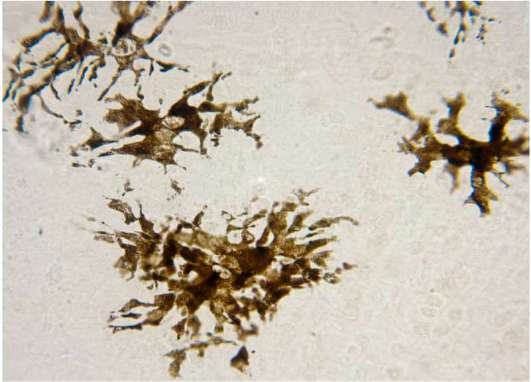
Рис. 8.7. Пигментоциты в коже (микрофотография). Препарат не окрашен
выявляются палочковидные и нитевидные митохондрии с плотно расположенными кристами. На периферии клетки встречаются многочисленные пиноцитозные пузырьки.
Адипоциты бурой жировой ткани отличаются мелкими липидными включениями и митохондриями, расположенными вокруг ядра. Адипоциты обладают большой способностью к метаболизму. Как количество жировых включений в адипоцитах, так и число самих жировых клеток в рыхлой соединительной ткани подвержено значительным колебаниям.
Функции адипоцитов - трофическая, связанная с обеспечением энергии и резерва воды в организме, а также участие в процессе терморегуляции.
Новые жировые клетки в соединительной ткани взрослого организма могут развиваться при усиленном питании из адвентициальных (камбиальных) клеток, прилегающих к кровеносным капиллярам. При этом в цитоплазме клеток появляются сначала мелкие капельки жира, которые, увеличиваясь в размере, постепенно сливаются в более крупные капли. По мере увеличения жировой капли эндоплазматическая сеть и комплекс Гольджи редуцируются, а ядро сдавливается, уплощается и оттесняется на периферию клетки.
Скопления жировых клеток, встречающихся во многих органах, называют жировой тканью.
Адвентициальные клетки. Это малодифференцированные (камбиальные) клетки, сопровождающие мелкие кровеносные сосуды. Они имеют уплощенную или веретенообразную форму со слабобазофильной цитоплазмой, овальным ядром и небольшим числом органелл. Эти клетки путем дивергентной дифференцировки дают начало различным клеточным дифферонам (фибробластическому, миофибробластическому, адипоцитарному и др.).
Перициты - клетки, окружающие кровеносные капилляры и входящие в состав их стенки (см. главу 13).
Пигментоциты (пигментные клетки, меланоциты). Эти клетки содержат в своей цитоплазме пигмент меланин. Их много в родимых пятнах, а также в соединительной ткани людей черной и желтой рас. Пигментоциты имеют короткие, непостоянной формы отростки (рис. 8.7), большое количество меланосом (гранул меланина) размером 15-25 нм и рибосом. Часть мелано-сом из меланоцитов мигрируют в кератиноциты шиповатого и базального слоев эпидермиса.
В цитоплазме меланоцитов содержатся также биологически активные амины, которые могут принимать участие вместе с тучными клетками в регуляции тонуса стенок сосудов (см. главу 14).
Меланоциты только формально относятся к соединительной ткани, так как располагаются в ней. Что касается их происхождения, то доказано образование этих клеток из стволовых клеток в составе нервных гребней, а не из мезенхимы. Клетки соединительной ткани функционально связаны в единую систему благодаря многочисленным факторам взаимодействия, особенно в процессах воспаления и посттравматической регенерации, при нарушении водно-солевого режима организма и др.
Межклеточное вещество
Межклеточное вещество, или матрикс (substantia intercellularis), соединительной ткани состоит из коллагеновых и эластических волокон, а также из основного (аморфного) вещества. Межклеточное вещество как у зародышей, так и у взрослых образуется, с одной стороны, путем секреции, осуществляемой соединительнотканными клетками, а с другой - из плазмы крови, поступающей в межклеточные пространства.
У зародышей человека образование межклеточного вещества происходит, начиная с 1-2-го мес внутриутробного развития. В течение жизни межклеточное вещество постоянно обновляется.
Коллагеновые структуры, входящие в состав соединительных тканей организмов человека и животных, являются наиболее представительными ее компонентами, образующими сложную организационную иерархию. Основу всей группы коллагеновых структур составляет волок-нис тый белок - коллаген, который определяет свойства коллагеновых структур.
Коллагеновые волокна (fibrae collagenae) в составе разных видов соединительной ткани определяют их прочность. В рыхлой неоформленной соединительной ткани они располагаются в различных направлениях в виде волнообразно изогнутых, спиралевидно скрученных, округлых или уплощенных в сечении тяжей толщиной 1-3 мкм и более. Длина их различна. Внутренняя структура коллагенового волокна определяется фибриллярным белком - коллагеном, который синтезируется на рибосомах гранулярной эндоплазматической сети фибробластов.
Различают около 20 типов коллагена, отличающихся молекулярной организацией, органной и тканевой принадлежностью.

Рис. 8.8. Биосинтез коллагена и фибриллогенез
В организме человека наиболее распространены следующие: коллаген I типа встречается главным образом в соединительной ткани кожи, сухожилиях, кости, роговице глаза, склере, стенке артерий; коллаген II типа входит в состав гиалиновых и фиброзных хрящей, стекловидного тела, роговицы; коллаген III типа находится в дерме кожи плода, в стенках крупных кровеносных сосудов, в ретикулярных волокнах органов кроветворения; IV типа - в базальных мембранах, капсуле хрусталика; V тип коллагена присутствует в хорионе, амнионе, эндомизии, пери-мизии, коже, вокруг клеток (фибробластов, эндотелиальных, гладкомышечных), синтезирующих коллаген.
Молекулы коллагена имеют длину около 280 нм и ширину 1,4 нм. Они построены из триплетов - трех полипептидных цепочек (α-цепочки) предшественника коллагена - проколлагена, свивающихся еще в клетке в единую спираль. Это первый, молекулярный, уровень организации коллагеново-го волокна. Проколлаген секретируется в межклеточное вещество (рис. 8.8).
Второй, надмолекулярный, уровень - внеклеточной организации колла-генового волокна - представляют собой агрегированные в длину и поперечно связанные с помощью водородных связей молекулы тропоколлаге-на, образующиеся путем отщепления концевых пептидов проколлагена. Сначала образуются протофибриллы, а 5-6 протофибрилл, скрепленных между собой боковыми связями, составляют микрофибриллы толщиной около 5 нм.
При участии гликозаминогликанов, также секретируемых фибробласта-ми, формируется третий, фибриллярный, уровень организации коллагенового волокна. Коллагеновые фибриллы представляют собой поперечно исчерчен-

Рис. 8.9. Коллагеновая фибрилла (препарат Н. П. Омельяненко). П - период. Электронная микрофотография. Увеличение 200 000
ные структуры толщиной в среднем 20-100 нм. Период повторяемости темных и светлых участков 64-67 нм (рис. 8.9, 8.10). Каждая молекула коллагена в параллельных рядах, как полагают, смещена относительно соседней цепи на четверть длины, что служит причиной чередования темных и светлых полос. В темных полосах под электронным микроскопом видны вторичные тонкие поперечные линии, обусловленные расположением полярных аминокислот в молекулах коллагена.
Четвертый, волоконный, уровень организации. Коллагеновое волокно, образующееся путем агрегации фибрилл, имеет толщину 1-10 мкм (в зависимости от топографии). В него входит различное количество фибрилл - от единичных до нескольких десятков. Волокна могут складываться в пучки толщиной до 150 мкм.
Коллагеновые волокна отличаются малой растяжимостью и большой прочностью на разрыв. В воде толщина сухожилия в результате набухания увеличивается на 50 %, а в разбавленных кислотах и щелочах - в 10 раз, но при этом волокно укорачивается на 30 %. Способность к набуханию больше выражена у молодых волокон. При термической обработке в воде коллаге-новые волокна образуют клейкое вещество (греч. kolla - клей), что и дало название этим волокнам.
Разновидностью коллагеновых волокон являются ретикулярные и прекол-лагеновые волокна. Последние представляют собой начальную форму образования коллагеновых волокон в эмбриогенезе и при регенерации. В их состав входят коллаген III типа и повышенное количество углеводов, которые синтезируются ретикулярными клетками органов кроветворения. Они образуют трехмерную сеть - ретикулум, что и обусловило их название.
Эластические волокна. Наличие эластических волокон (fibra elasticae) в соединительной ткани определяет ее эластичность и растяжимость. По прочности эластические волокна уступают коллагеновым. Форма поперечного разреза волокон округлая и уплощенная. В рыхлой соединительн
|
|
|
