 |


Синтез и распад гликогена.
|
|
|
|
Синтез гликогена протекает не во всех тканях, а только в печени, мышцах и в лейкоцитах.
После образования глюкозо-6-фосфата (гексокиназная реакция) происходит внутримолекулярный перенос остатка фосфорной кислоты из 6-го положения в 1-е. При этом образуется глюкозо-1-фосфат:

После изомеризации глюкозо-6-фосфата в глюкозо-1-фосфат протекает дополнительная активация глюкозного фрагмента. При этом расходуется 1 молекула УТФ, что эквивалентно расходованию 1-й молекулы АТФ. В результате образуется активированная форма - УДФ-глюкоза:
Затем с УДФ глюкозный остаток переносится на молекулу гликогена. Удлинение цепи гликогена катализирует фермент гликогенсинтетаза. Таким образом, цепь гликогена становится на 1 глюкозный фрагмент длиннее. Гликоген, в отличие от растительного крахмала, более сильно разветвлен. Для формирования ответвлений существует специальный фермент, который называется "гликогенветвящий фермент" (стр.242 учебника).
Молекула гликогена синтезируется не с "нуля", а происходит постепенное удлинение уже имеющегося кусочка цепи: "затравки". И при распаде гликогена никогда не происходит полного разрушения его молекул.
Для включения одного остатка глюкозы в молекулу гликогена клетка расходует 2 молекулы АТФ. При распаде гликогена эта АТФ не регенерирует, а освобождается только Фн (неорганический фосфат).
Ключевым ферментом синтеза гликогена является гликогенсинтаза. Это "пункт вторичного контроля". Ее Vmax больше, чем у гексокиназы, но меньше, чем у любого другого фермента на пути синтеза гликогена.
Регуляция гликогенсинтазы: она активируется избытком глюкозо-6-фосфата. Поэтому если глюкозо-6-фосфат другими путями утилизируется медленно, то возрастание его концентрации приводит к увеличению скорости синтеза гликогена.
|
|
|
Реакция, катализируемая гликогенсинтазой, необратима.
В определенных условиях гликоген способен распадаться. Для этого существует свой обходной обратный путь. Его ключевым ферментом является гликогенфосфорилаза (фосфорилаза). Этот фермент расщепляет молекулу гликогена с участием Фн до глюкозо-1-фосфата и гликогена, укороченного на один глюкозный фрагмент: (С6Н10О5)n + H3PO4 ----> (C6H10O5)n-1 + глюкозо-1-фосфат
Фосфорилаза - ключевой (то есть лимитирующий и регуляторный) фермент распада гликогена.
Регуляция гликогенфосфорилазы: угнетается избытком АТФ, активируется избытком АДФ.
Г Б Ф - П У Т Ь.
(ГЕКСОЗОБИСФОСФАТНЫЙ ПУТЬ РАСПАДА УГЛЕВОДОВ)
БИОЛОГИЧЕСКОЕ ЗНАЧЕНИЕ ГБФ-ПУТИ.
1. Это главный путь распада углеводов до конечных продуктов. Во многих клетках это - единственный путь. Так распадается 70-75% глюкозы, которая поступает в клетку.
2. Только ГБФ-путь дает клетке энергию в виде АТФ. Это основной источник получения энергии в клетке.
3. Это самый длинный путь распада углеводов.
ГБФ-путь делим на 3 этапа.
1-й этап протекает в цитоплазме, дает 8 молекул АТФ при распаде 1 молекулы глюкозы или 9АТФ при распаде одного глюкозного фрагмента гликогена. Заканчивается образованием 2-х молекул пирувата (ПВК).
2-й и 3-й этапы - (исключительно аэробные!) в митохондриях с обязательным участием кислорода, дают 30 АТФ в расчете на одну молекулу глюкозы.
2-й этап ГБФ-пути называется "окислительное декарбоксилирование пирувата" и катализируется пируватдегидрогеназным комплексом (смотрите лекции "Биологическое окисление" - удлиненная цепь митохондриального окисления). На 2-м этапе от молекулы ПВК отнимаются два атома водорода, и пируват превращается в Ацетил-кофермент А (АцКоА), одновременно происходит отщепление СО2. Два атома водорода идут на НАД, а затем по цепи митохондриального окисления передаются на О2 с образованием Н2О и 3 молекул АТФ. Поэтому в расчете на одну молекулу исходной глюкозы 2-й этап дает 6 АТФ.
|
|
|
В 3-й этап вступает молекула АцетилКоА, который образуется в результате 2-го этапа. Этот 3-й этап называется циклом трикарбоновых кислот (ЦТК) (смотрите лекции “Митохондриальное окисление”). В этом цикле АцКоА полностью расщепляется до СО2 и Н2О. При этом образуется 12 АТФ в расчете на молекулу АцКоА, вступившую в цикл. Если рассчитать на 1 молекулу глюкозы, то на 3-м этапе образуется 24 АТФ.
1-й этап проходит 10 промежуточных стадий. В ходе первой части этого этапа молекула глюкозы расщепляется пополам до 2-х молекул фосфоглицеринового альдегида (ФГА).
ОСОБЕННОСТИ ПЕРВОЙ ЧАСТИ 1-ГО ЭТАПА:
Гексокиназа (ГК) работает, чтобы ослабить прочную молекулу глюкозы:

2-я реакция - изомеризации:

На 3-й стадии фруктозо-6-фосфат еще более ослабляется фосфофруктокиназой (ФФК) и образуется фруктозо-1,6-бисфосфат:

|
Фосфофруктокиназа - это ключевой фермент ГБФ-пути. Он является "пунктом вторичного контроля". Vmax ФФК больше, чем Vmax ГК. Поэтому, когда глюкозы поступает много, ГК лимитирует скорость всего ГБФ-пути.
Избыток АТФ и избыток цитрата сильно ингибируют ФФК. В этих условиях лимитирующим ферментом ГБФ-пути вместо гексокиназы становится ФФК. Из-за угнетения ФФК накапливаются глюкозо-6-фосфат (Г-6-Ф) и фруктозо-6-фосфат(Ф-6-Ф). Г-6-Ф ингибирует гексокиназу, уменьшая утилизацию глюкозы клеткой и одновременно активирует гликогенсинтетазу.
Если нет избытка АТФ и цитрата, а есть избыток АДФ, то АДФ активирует ФФК, и тогда скорость всего ГДФ-пути лимитируется опять гексокиназой.
В результате фосфофруктокиназной реакции молекула фруктозо-1,6-бисфосфата дестабилизируется (ослабляется) настолько, что сразу распадается на 2 триозы при участии фермента альдолазы (4-я реакция):
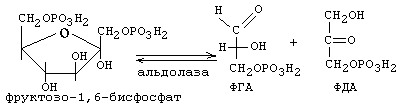
5-я реакция:
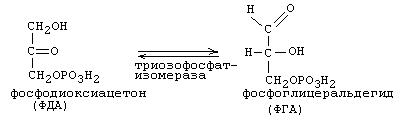
В следующую (шестую) реакцию ГБФ-пути вступает только ФГА. В результате уменьшается его концентрация и равновесие 5-й реакции сдвигается в сторону образования ФГА. Постепенно весь ФДА переходит в ФГА, и поэтому количество АТФ, синтезировавшееся в последующих реакциях ГБФ-пути, мы учитываем в расчете на 2 молекулы ФГА и других промежуточных метаболитов, которые из него образуются.
|
|
|
В 1-й части 1-ого этапа (от глюкозы до ФГА) расходуется 2 молекулы АТФ: одна - в гексокиназной реакции, другая - в фосфофруктокиназной (3-я реакция первого этапа ГБФ-пути). 2-я часть 1-го этапа начинается с окисления ФГА до ФГК (фосфоглицериновой кислоты) в 6-й реакции.

|
Эта реакция катализируется ферментом "глицеральдегидфосфатдегидрогеназа". Отщепляемый водород передается на НАД с образованием НАДН2. Энергии, которая выделяется при этом окислении, хватает и на то, чтобы одновременно обеспечить присоединение фосфата к альдегидной группе. Присоединяется фосфат макроэргической связью. В результате образуется 1,3-дифосфоглицериновая кислота (1,3-бисфосфоглицерат).
7-я реакция: субстратное фосфорилирование.
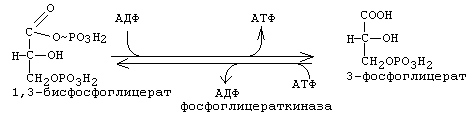
Фосфат с макроэргической связью передается на АДФ с образованием АТФ. В результате 7-й стадии в молекуле фосфоглицериновой кислоты остается 1 остаток фосфорной кислоты.
8-я реакция: Фосфат переносится из 3-го во второе положение и образуется 2-фосфоглицериновая кислота.

9-я реакция:

|
От 2-фосфоглицериновой кислоты отнимается Н2О. Это приводит к перераспределению молекулярной энергии. В результате на фосфате во втором положении накапливается энергия и связь становится макроэргической. Получается фосфоенолпируват(ФЕП).
10-я реакция: Субстратное фосфорилирование. Фосфат переносится на АДФ с образованием АТФ. ФЕП переходит в ПВК (пировиноградную кислоту).

На этом 1-й этап ГДФ-пути заканчивается, ПВК уходит в митохондрию и вступает во второй этап ГДФ-пути.
Итоги 1-го этапа: 10 реакций, из которых первая, третья и десятая реакции необратимы. Сначала расходуется 2 АТФ на 1 молекулу глюкозы. Потом окисляется ФГА. Энергия реализуется в ходе 2-х реакций субстратного фосфорилирования: в каждой из них образуется по 2 АТФ. Следовательно, на каждую молекулу глюкозы (на 2 молекулы ФГА) получается 4 АТФ путем субстратного фосфорилирования.
|
|
|
Суммарно все 10 стадий можно описать следующем уравнением:
С6Н12О6 + 2Н3РО4 + 2АДФ + 2НАД -----> 2С3Н4О3 + 2АТФ + 2Н2О + 2НАДН2
(ПВК)
НАДН2 по системе митохондриального окисления(МтО) передает водород на кислород воздуха с образованием Н2О и 3 АТФ, но 1-й этап протекает в цитоплазме и НАДН2 не может проходить через мембрану митохондрий. Существуют челночные механизмы, обеспечивающие этот переход НАДН2 через митохондриальную мембрану - малат-аспартатный челнок и глицерофосфатный челнок (смотрите лекции "Биологическое окисление".
В расчете на одну молекулу глюкозы образуется 2НАДН2.

|
В дополнение к 2 АТФ, получаемым на 1-м этапе путем субстратного фосфорилирования, образуется еще 6 АТФ с участием кислорода, итого - 8 молекул АТФ. Столько АТФ образуется в расчете на каждую расщепленную до ПВК молекулу глюкозы в ходе первого этапа ГБФ-пути.
Если эти 8 АТФ добавить к 30 молекулам АТФ, которые образуются на 2-м и 3-м этапах, то суммарный энергетический итог всего ГБФ-пути составит 38 АТФ на каждую молекулу глюкозы, расщепленную до СО2 и Н2О. В этих 38 АТФ заключено 65 процентов энергии, которая выделилась бы при сжигании глюкозы на воздухе. Это доказывает очень высокую эффективность работы ГБФ-пути.
Из 38 АТФ основная их часть образуется на 2-м и 3-м этапах. Каждый из этих этапов абсолютно необратим и требует обязательного участия кислорода, так как окислительные стадии этих этапов сопряжены с митохондриальным окислением (без него невозможны). Весь ГБФ-путь от глюкозы или гликогена до СО2 и Н2О называют: АЭРОБНЫЙ РАСПАД УГЛЕВОДОВ.
Ключевые ферменты первого этапа ГБФ-пути: ГЕКСОКИНАЗА и ФОСФОФРУКТОКИНАЗА.
Еще одно ключевое звено находится в ЦТК (3-й этап ГБФ-пути). Ключевое звено на 3-м этапе необходимо потому, что АцКоА, вступающий в ЦТК, образуется не только из углеводов, но и из жиров и аминокислот. Следовательно, ЦТК - это конечный "котёл" для сжигания ацетильных остатков, образующихся из углеводов, жиров и белков. ЦТК объединяет все метаболиты, образующиеся при распаде углеводов, жиров и белков.
Ключевые ферменты ЦТК: цитратсинтетаза и изоцитратдегидрогеназа. Оба фермента угнетаются избытком АТФ и избытком НАДН2. Изоцитратдегидрогеназа активируется избытком АДФ. АТФ ингибирует эти ферменты по-разному: изоцитратдегидрогеназа ингибируется АТФ намного сильнее, чем цитратсинтаза. Поэтому при избытке АТФ накапливаются промежуточные продукты: цитрат и изоцитрат. В этих условиях цитрат может выходить в цитоплазму по градиенту концентраций.
2-й и 3-й этапы ГБФ-пути протекают в митохондриях, а 1-й - в цитоплазме.
|
|
|
1-й этап отделен от 2-го и 3-го этапов митохондриальной мембраной.
Поэтому 1-й этап может выполнять свои особенные функции. Эти функции
связаны с двумя особенностями 1-го этапа.
|
|
|
