 |


Развитие аксонов ганглиозных клеток.
|
|
|
|
Радужка
Развитие радужной оболочки во многом зависит от полного и правильного закрытия эмбриональной щели, а также от своевременного образования передней части «сосудистой сумки хрусталика» (tunica vasculosa lends) [6, 14, 49, 139]. Сосудистые каналы этой эмбриональной структуры первоначально возникают на шестой неделе развития (размер эмбриона равен 17— 18 мм) в виде кольцевидного сосуда, окружающего край зрительного бокала. Тяжи мезен-химных клеток продвигаются к передней поверхности хрусталика. Эта мезенхимная ткань представляет собой вторую волну миграции клеток нервного гребня, участвующих в образовании передней камеры глаза и, в конечном счете, дифференцирующихся в строму радужной оболочки (рис. 5.7.3—5.7.5).
Сосудистая сеть радужки. Задние длинные ресничные артерии с носовой и височной сторон ресничного тела разветвляются и их ветви впадают в сосуды, лежащие по периферии «сосудистой сумки хрусталика». Таким образом, образуется большой артериальный круг кровообращения радужки. Позже, большой артериальный круг получает ветви от сплетения передних ресничных артерий. Сосудистые ветви задних длинных ресничных артерий и большого артериального круга кровоснабжают зрачковую мембрану.
После того как к концу третьего месяца эмбрионального развития сформировались ресничные складки (будущие ресничные отростки), края обеих стенок зрительного бокала быстро растут вперед.
На стадии развития, при которой эмбрион достигает 48—50 мм, от зрительного бокала отделяется периферическая часть «сосудистой сумки хрусталика», которая становится сосудистой сетью зрачковой мембраны. Таким образом, к концу четвертого месяца сосудистая система радужки состоит из двух слоев: сзади — сосуды «сосудистой сумки хрусталика», а спереди — сосуды зрачковой мембраны.
|
|
|
На протяжении пятого месяца развития ветви, отходящие от длинных ресничных артерий,

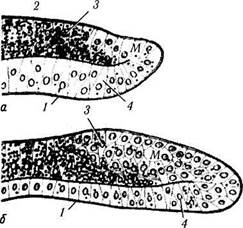
Рис. 5.7.5. Развитие радужной оболочки и ресничного тела (8-й месяц развития):
а — радужная оболочка и угол передней камеры; б — область зубчатой линии. Четко определяется угол передней камеры, корень радужки и ресничное тело (/ — зубчатая линия; 2 — сенсорная часть сетчатки; 3 — ресничное тело; 4 — радужка; 5 — угол передней камеры; 6 — роговая оболочка; 7 — склера)
достигают мезенхимы в средней части радужки и образуют сосудистые арки.
Мезенхимные производные радужки дифференцируются раньше, чем производные нейро-эктодермы. На передней поверхности радужки мезенхимные клетки образуют однослойную выстилку, отграничивающую переднюю камеру глаза. В последующем они участвуют в образовании переднего пограничного слоя радужки. Межклеточных контактов между клетками нет, а видны лишь широкие промежутки, выполненные цитоплазматическими отростками.
На последующих этапах в строме радужки появляются пигментированные меланоциты (9 месяцев эмбриогенеза) [153, 105]. Степень пигментации определяется расовой принадлежностью индивидуума и достигает максимума после рождения.
Часть мезенхимных клеток стромы превращается в фибробластоподобные клетки, секре-тирующие коллагеновые фибриллы и межклеточное вещество.
Сфинктер и дилятатор (рис. 5.7.6, 5.7.7). Дальнейший рост и дифференциация двухслойного нейроэктодермального слоя зрительного бокала приводит к образованию мышц радужки — дилятатора и сфинктера.
Начало формирования сфинктера сводится к появлению выпячиваний на базальной поверхности клеток переднего эпителиального слоя.
Глава 5. РАЗВИТИЕ ГЛАЗНОГО ЯБЛОКА

|

|
| Д««ИИЖЫМГ»МЯ |
|
|
|
Рис. 5.7.6. Схемы развития сфинктера радужной оболочки (по Ruprecht, Wulle, 1973):
а — 3-й месяц эмбрионального развития; б— 4-й месяц эмбрионального развития; в — 5-й месяц эмбрионального развития; г—8-й месяц эмбрионального развития (/ — базальная мембрана; 2 — будущая строма радужной оболочки; 3 — наружный слой эпителия;
4 — внутренний слой эпителия; М — сфинктер)

|
К 3-му месяцу клетки начинают синтезировать базальную мембрану, элементы цитоскелета, характерные для гладкомышечных клеток, а на протяжении пятого месяца и миофибриллы [263]. Даже при появлении миофиламентов и межклеточных контактов мышца не отделяется от пигментного эпителия. На шестом месяце развития в сформированные мышечные пучки проникают соединительнотканные тяжи и кровеносные сосуды. При этом отмечается начало отделения мышцы от эпителия. Завершается процесс погружения мышцы в строму лишь на 8-м месяце.
|
Признаков развития дилятатора нет до шестого месяца. Первый признак развития мышцы характеризуется появлением в эпителиальных клетках, лежащих вблизи шпоры Михеля (Michel), нежных миофибрилл. Миоэпителиаль-ные клетки веретеновидной формы. Четко видна тонкая базальная мембрана, расположенная между мышечными клетками и стромой радужки [254]. Миоэпителиальные клетки продолжают развиваться и после рождения. Дилятатор, в отличие сфинктера, не отделяется от эпителия и не окружен стромой.
| Рис. 5.7.7. Стадии развития сфинктера радужной оболочки из периферической части края глазного бокала (по Ruprecht, Wulle, 1973): а —12-я неделя эмбрионального развития; б — 7-й месяц эмбрионального развития (/ — наружный слой глазного бокала; 2 — внутренний слой глазного бокала; 3 —место формирования будущего сфинктера; 4 — шпора Михеля; 5 — мезенхима, формирующая зрачковую (иридопупиллярную) мембрану; 6 —строма роговой оболочки; 7 —эндотелий роговой оболочки; 8 —«сосудистая сумка хрусталика»; 9 —капсула хрусталика) |
Пигментный эпителий. Слой пигментного эпителия начинается в области зрачкового края радужки и прерывается у его корня. Пигментация эпителиальных клеток обнаруживается лишь к концу седьмого месяца. В этот промежуток времени на задней поверхности радужки, обращенной к задней камере глаза, в эпителиальных клетках появляются многочисленные складки. На протяжении эмбрионального развития сила сцепления между двумя слоями эпителиальных клеток незначительная, особенно в области зрачка. В результате этого между двумя эпителиальными слоями существует циркулярный синус (краевой синус фон Зили (von Szily)) (рис. 5.7.4).
|
|
|
Развитие увеального тракта
 «Воротничок». Возникновение «воротничка» в строме радужки связано с тем, что в эмбриогенезе на сфинктере располагаются петли сосудов зрачковой мембраны. На протяжении шестого месяца наступает регрессия центральных участков «сосудистой сумки хрусталика». Сосуды атрофируются, и при этом прекращается кровообращение. Сохраняются лишь сосуды, которые на последующих стадиях образуют малый круг кровообращения радужки, расположенный по краю сфинктера. Эта линия и является границей «воротничка». К девятому месяцу поверхностно расположенные сосуды также атрофируются. «Воротничок» у новорожденного располагается ближе к зрачку, чем у взрослых.
«Воротничок». Возникновение «воротничка» в строме радужки связано с тем, что в эмбриогенезе на сфинктере располагаются петли сосудов зрачковой мембраны. На протяжении шестого месяца наступает регрессия центральных участков «сосудистой сумки хрусталика». Сосуды атрофируются, и при этом прекращается кровообращение. Сохраняются лишь сосуды, которые на последующих стадиях образуют малый круг кровообращения радужки, расположенный по краю сфинктера. Эта линия и является границей «воротничка». К девятому месяцу поверхностно расположенные сосуды также атрофируются. «Воротничок» у новорожденного располагается ближе к зрачку, чем у взрослых.
Адренергическая и холинергическая иннервация стромы радужки обнаруживается только после развития мышц при размере эмбриона 200—300 мм (7,5 мес.) [132, 168].
5.8. РАЗВИТИЕ СЕТЧАТОЙ ОБОЛОЧКИ
Особенности развития сетчатой оболочки описаны в многочисленных исследованиях, которые продолжаются и по сей день [6, 14, 42, 49, 139]. Если первоначально эти работы были направлены на изучение чисто морфологичес-
ких процессов эмбриогенеза сетчатки, то в последнее время основная часть работ посвящена изучению тонких механизмов регуляции роста и дифференциации отдельных структур биологически активными веществами, синтезируемыми в процессе развития. Этому способствовало совершенстование морфологических и создание новых иммуногистохимических и цитогенетичес-ких методов исследования.
Морфологические проявления дифференциации нейроэктодермального слоя в направлении сенсорной части сетчатки начинаются довольно рано. Основные этапы этого процесса представлены на рис. 5.8.1—5.8.3.
|
|
|
На четвертной неделе эмбрионального развития (размер эмбриона равен 4 мм), как раз в момент завершения процесса образования зрительного пузырька, клетки внутреннего слоя зрительного пузырька митотически делятся, в результате чего образуется три или четыре ряда компактно расположенных клеток [198]. Со стороны будущей полости стекловидного тела клетки сетчатки прилежат к тонкой ба-зальной мембране. По бокам зрительного пузырька отростки зародышевых клеток сетчатки простираются в окружающую мезенхиму, расположенную под наружной эктодермой.
К 32-му дню (эмбрион 7 мм) сетчатка состоит из пяти или шести рядов нейроэпите-лиальных клеток. Ядра этих клеток отделены
 4—5-я недели развития
4—5-я недели развития
6-я неделя —3-й месяц развития
3—7-й месяцы развития
Взрослые
 Поверхность краевого (маргинального) слоя
Поверхность краевого (маргинального) слоя
Внутренняя пограничная мембрана
Маргинальный слой свободный от ядер
Поверхностная часть маргинального слоя
Слой нервных волокон
Слой нервных волокон
|

|

|
|
Нейроэпителий
Внутренний слой ней-робластов
Переходный слой Чи-вица
Наружный слой ней-робластов
Ганглиозные клетки Амакриновые клетки
Ядра мюллеровских клеток
Биполярные клетки
Горизонтальные клетки
Ядра палочек и колбочек
Ганглиозные клетки
Внутренний плексиформ-ный слой
Внутренний ядрный слой
Наружный плексиформ-ный слой
Наружный ядерный слой
Наружная пограничная мембрана
Реснички
Реснички
Примитивные палочки и колбочки
Палочки и колбочки
 Рис. 5.8.1. Особенности происхождения нейронов сетчатой оболочки (по Duke-Elder, 1963)
Рис. 5.8.1. Особенности происхождения нейронов сетчатой оболочки (по Duke-Elder, 1963)
Глава 5. РАЗВИТИЕ ГЛАЗНОГО ЯБЛОКА

|
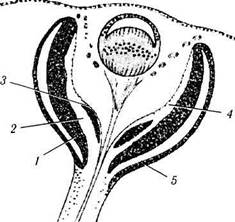
Рис. 5.8.2. Схема начального этапа развития сенсорной части сетчатки и ее пигментного эпителия:
/ — наружный слой нейробластов; 2 — слой Чивица; 3 — внутренний слой нейробластов; 4 — внутренняя краевая (маргинальная) зона; 5 — пигментный эпителий

|
|
| 11 |
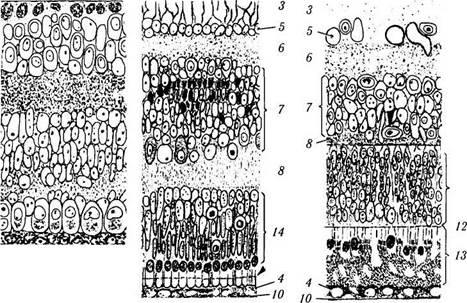
Рис. 5.8.3. Схема дифференциации слоев сетчатой оболочки во время эмбрионального развития:
а — 2,5 месяца эмбрионального развития. Переходный слой Чивица, разделяющий на более ранних этапах эмбриогенеза наружный и внутренний нейробластные слои, постепенно исчезает в результате перемещения в него тел нейронов и развития их отростков. Клетки, расположенные вблизи стекловидного тела дифференцируются в ганглиозные клетки. Клетки, расположенные наружу (звездочки), дифференцируются в амакриновые и мюллеровские клетки. Наружный слой нейробластов дифференцируется в фоторецепторные, биполярные и горизонтальные нейроны; б — 4,5 месяца эмбрионального развития. Ганглиозные клетки располагаются в несколько слоев. Внутренний плекси-формный слой состит из отростков биполярных, ганглиозных и амакриновных клеток. Появляются синаптические образования. В наружном ядерном слое клетки ориентируются своими апикальными поверхностями по направлению будущего пигментного эпителия. В наружном плексиформном слое определяются примитивные синаптические образования между дендритами биполярных клеток и ножками колбочек. Наружные членики фоторецепторов пока не выявляются; в — 5,5 месяца эмбрионального развития. Ганглиозные клетки формируют слой, состоящий из одного или двух рядов клеток. В состав внутреннего ядерного слоя входят амакриновые клетки с большими бледными ядрами и мюллеровские клетки, отличающиеся темным ядром. Эти два
|
|
|
типа клеток исходят из внутреннего нейробластого слоя. Также определяются небольшие биполярные клетки и крупные горизонтальные клетки. Последние два типа клеток дифференцируются из наружного нейробластного слоя, как и фоторецепторы. В наружном плексиформном слое обнаруживается линейное расположение синапсов между биполярными и фоторецепторными клетками. Наружный ядерный слой состоит из 6—7 слоев ядер клеток. Отмечается рост наружных члеников фоторецепторов в направлении пигментного эпителия (стрелка). Гибель клеточных элементов путем апоптоза обозначена темно окрашенными клетками; г — строение сетчатки на момент рождения. Сетчатка по строению приближается к сетчатке взрослого. Наружный плексиформный слой тоньше, чем у взрослых. Синапсы полностью сформированы. Наружные членики колбочек входят в контакт с пигментным эпителием (/ — внутренний нейробласт-ный слой; 2 — наружный нейробластный слой; 3 — слой нервных волокон; 4 — пигментный эпителий; 5 — слой ганглиозных клеток; 6 — внутренний плексиформный слой; 7— внутренний ядерный слой; 8 — наружный плексиформный слой; 9 — слой фоторецепторов; 10 — мембрана Бруха; // — слой расположения ядер фоторецепторов; 12 — наружная пограничная мембрана; 13 —палочки и колбочки; 14 — наружный ядерный слой. Стрелками указаны кровеносные сосуды сетчатки)
Развитие сетчатой оболочки
 от наружных 2/з зрительного бокала слоем, лишенным ядер и называемым внутренним краевым (маргинальным) слоем [198]. Подобный вид сетчатка сохраняет до седьмой недели эмбрионального развития.
от наружных 2/з зрительного бокала слоем, лишенным ядер и называемым внутренним краевым (маргинальным) слоем [198]. Подобный вид сетчатка сохраняет до седьмой недели эмбрионального развития.
Клетки наиболее наружного слоя ядерной зоны (называемые эпендимным, герминативным или пролиферативным слоями) обладают короткими цитоплазматическими отростками, обращенными в полость между двумя нейроэпите-лиальными слоями. Позже из них развиваются реснички, контактирующие с апикальной поверхностью клеток пигментного эпителия [104]. Эти структуры исчезают к седьмой неделе, а к четвертому месяцу замещаются предшественниками наружных члеников фоторецепторов.
Необходимо отметить, что дифференциация сенсорной части сетчатки у большинства животных и приматов начинается в ее заднем отделе и продолжается кпереди [139, 189, 192, 200]. Причем эта дифференциация начинается именно в области будущей фовеолы [131, 167, 188]. В противоположность сенсорной части сетчатки пигментный эпителий сетчатки начинает дифференцироваться в обратном направлении [24, 25].
В момент начала дифференциации макуляр-ной области она представляет собой лишь еле заметную точку [184].
На этом этапе развития митотическая активность нейроэпителиальных клеток характерна для всей поверхности будущей сетчатки, но
уже к 14-й неделе митозы исчезают в ее центральной части. Именно в этой, четко очерченной, области (площадь примерно 2%) появляются признаки начала дифференциации колбочек (предположительно область фовеа).
По мере дальнейшей дифференциации структур сетчатки количество митозов все более уменьшается так, что к 24 неделям делящиеся клетки определяются только в 62,5% поверхности сетчатки, причем только по периферии. К 30-й неделе митотическая активность полностью прекращается и, в первую очередь, в области будущей фовеолы [42, 79, 269, 189, 200].
Тем не менее площадь поверхности сетчатки все же увеличивается, причем скорость увеличения равна 10—15 мм2 в неделю. Эта скорость сохраняется на протяжении всего эмбрионального развития и в течение первых трех недель после рождения. Увеличение площади сетчатки в постмитотической стадии связано с дальнейшей дифференциацией клеток [184, 185].
Организация нейроэпителия. К 4—5-й неделям эмбриогенеза (эмбрион 12 мм) нейро-эпителий состоит из двух неиробластных слоев: внутреннего, обращенного к стекловидному телу, и наружного, граничащего с пигментным эпителием сетчатки [6, 14, 49, 139]. Наружный слой нейробластов образуется в результате миграции будущих ганглиозных клеток и клеток Мюллера из наружного нейроэпителиаль-ного слоя (рис. 5.8.2—5.8.4). В результате это-
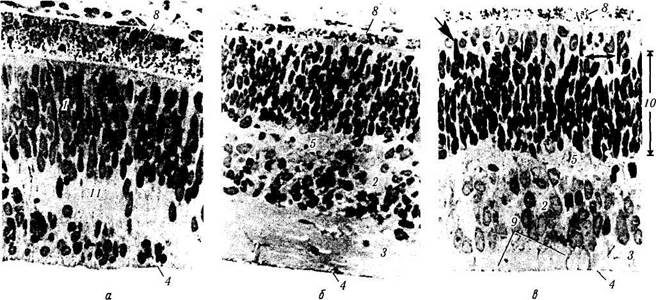
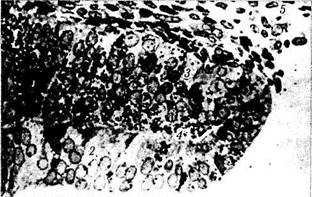
Рис. 5.8.4. Микроскопическая картина развития сенсорной части сетчатки и пигментного эпителия (по Spira,
Hellenberg, 1973):
а — эмбрион на седьмой неделе развития. Клетки наружного и внутреннего неиробластных слоев разделены безъядерным слоем Чивица. Четко определяются наружная и внутренняя пограничные мембраны (стрелки); б — эмбрион на десятой неделе развития. Слой Чивица превращается в дифинитивный внутренний плексиформныи слой; в — двенадцатая неделя развития. Видно, как отростки мюллеровских клеток разделяют на пучки слой нервных волокон (/ — наружный нейробластный слой; 2 — слой ганглиозных клеток; 3 — слой нервных волокон; 4 — внутренняя пограничная мембрана; 5 — внутренний плексиформныи слой; 6 — внутренний ядерный слой; 7 — колбочки (стрелкой указаны клетки, являющиеся предшественниками колбочек); 8 — пигментный эпителий;
9 — волокна клеток Мюллера; 10 — слой Чивица)
Глава 5. РАЗВИТИЕ ГЛАЗНОГО ЯБЛОКА
 го процесса остается область, состоящая только из клеточных отростков. Называется этот слой переходным слоем Чивица. В этот период времени на основе различий строения ядра можно различить три типа клеток [198]:
го процесса остается область, состоящая только из клеточных отростков. Называется этот слой переходным слоем Чивица. В этот период времени на основе различий строения ядра можно различить три типа клеток [198]:
1. Клетки с недифференцированными оваль
ными ядрами, компактным хроматином и не
сколькими ядрышками. Расположены эти клет
ки преимущественно в наружном слое нейро-
бластов.
2. Клетки с большими круглыми или оваль
ными ядрами с нежным кружевным хромати
ном. Эти клетки обнаруживаются, главным об
разом, во внутреннем слое. Они обладают ши
роким ободком цитоплазмы. Предполагают, что
эти клетки в будущем дифференцируются в
ганглиозные клетки.
3. Клетки с темными круглыми или оваль
ными ядрами. При этом они небольшого разме
ра и располагаются во всех слоях развиваю
щейся сетчатки. Наиболее многочисленны они
во внутренней половине внутреннего слоя ней-
робластов. Считают, что подобные клетки в
последующем превращаются в мюллеровские
клетки.
Слои сетчатки (рис. 5.8.2—5.8.4). На протяжении 9—12-й недель слой Чивица превращается в дефинитивный внутренний плекси-формный слой. При этом уже можно различить четыре горизонтальных слоя сетчатки [198]. Ганглиозные клетки теперь лежат отдельно, а их аксоны образуют пучки, окруженные отростками мюллеровских клеток.
К десятой неделе эмбрионального развития наиболее поверхностные клетки наружного неиробластного слоя начинают превращаться в колбочки. Боковые поверхности этих клеток соединяются при помощи межклеточных контактов, и образуется наружная пограничная мембрана [104]. Развивающиеся отростки мюллеровских клеток следуют за нейронами во внутренние и наружные слои сетчатки. Межклеточные контакты также обнаруживаются между наружной плазматической мембраной развивающихся фоторецепторов и внутренней мембраной клеток пигментного эпителия. Благодаря этим контактам полость зрительного пузырька полностью смыкается.
Между 14-й и 17-й неделями число ганглиоз-ных клеток быстро нарастает, а между 18-й и 30-й неделями скорость их пролиферации резко снижается [184, 185]. К концу беременности количество клеток существенно снижается, что связано с гибелью клеток путем апоптоза.
Ганглиозные клетки в центральных участках сетчатки первоначально небольшого размера (4—10 мкм в диаметре) и имеют веретенообразную форму. По мере дальнейшего развития объем их тела значительно увеличивается.
В последние годы благодаря использованию методов ауторадиографии показано, что различ-
ные классы ганглиозных клеток сетчатки, отличающиеся морфологически и функционально (см. Сетчатка), дифференцируются на различных стадиях развития. Об этом свидетельствует различное время выхода их из митотического цикла [193, 223, 269, 270]. Так, у кошки ганглиозные клетки среднего размера возникают перед большими, а маленькие — в конце процесса дифференциации. По всей видимости, указанная закономерность «вызревания» ганглиозных клеток обеспечивает ретинотопическую проекцию их на нейроны наружного коленчатого тела. Связано это с тем, что различного типа ганглиозные клетки преимущественно располагаются в различных частях сетчатой оболочки и при росте аксонов, естествено в разное время, достигают зрительного перекреста, где они либо перекрещиваются в соответствии с топографическим расположением, либо достигают нейронов наружного коленчатого тела без перекрещивания.
Амакриновые клетки, идентифицируемые благодаря наличию большого бледного круглого ядра, первоначально рассеяны вдоль внутренней границы наружного неиробластного слоя (14-я неделя, эмбрион 70 мм).
Колбочки продолжают увеличиваться. К концу 17-й недели появляются синапсы во внутреннем плексиформном слое. Формирование синапсов ножек колбочек отмечается в наружном плексиформном слое в конце третьего месяца. В этот же период времени образуются и синаптические ленты, полностью имеющие дифференцированное строение только перед рождением [104]. Таким образом, синаптические структуры развиваются раньше, чем наружные членики фоторецепторов.
К 18-й неделе (эмбрион 96 мм) наружная часть цитоплазмы колбочки принимает зернистый вид в результате накопления митохондрий и полисом. В этот период времени можно обнаружить базальное тельце ресничек. Мембрана фоторецепторов позже (5-й месяц) отдает цито-плазматические отростки цилиндрической формы по направлению клеток пигментного эпителия. Наружные членики фоторецепторов начинают дифференцироваться к 5-му месяцу, а формирование мембранных стопок обнаруживается на 7-м месяце эмбрионального развития [287] (рис. 5.8.5—5.8.7).
К 22-й неделе (размер эмбриона равен 120 мм) между фоторецепторами видны клетки с темными ядрами. Это тела клеток палочек. Их развивающиеся внутренние членики первоначально выступают за пределы наружной пограничной мембраны [104].
Биполярные клетки не обнаруживаются во внутреннем ядерном слое до 23-й недели, но уже к 25-й неделе их дендриты простираются на наружный плексиформный слой. Горизонтальные клетки, вероятно, развиваются в то же самое время [198].
Развитие сетчатой оболочки


|

|
|
|

|

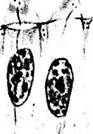
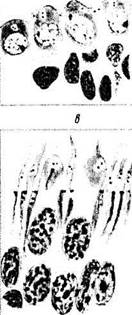
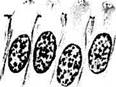
Рис. 5.8.5. Стадии формирования палочек и колбочек (по Bach, Seejelder):
|
| Са |
 а, б — эмбрион 65 мм. Колбочковые клетки в центральных участках сетчатки; в — эмбрион 80 мм. Колбочковые клетки в центральных участках сетчатки; г — эмбрион 345 мм. Колбочки в парацентральных участках сетчатки; <Э — эмбрион 345 мм. Палочки и колбочки в парацентральных участках сетчатки; е — эмбрион 420 мм. Колбочки центральных участков сетчатки
а, б — эмбрион 65 мм. Колбочковые клетки в центральных участках сетчатки; в — эмбрион 80 мм. Колбочковые клетки в центральных участках сетчатки; г — эмбрион 345 мм. Колбочки в парацентральных участках сетчатки; <Э — эмбрион 345 мм. Палочки и колбочки в парацентральных участках сетчатки; е — эмбрион 420 мм. Колбочки центральных участков сетчатки
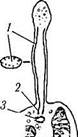
Рис. 5.8.7. Схема дифференциации наружного членика
палочек (б) из первичной реснички (а) (по Робертис
и соавт., 1967):
1 — нити, образующие соединительную ресничку; 2, 3 — цент-риоли, 4 — митохондрии; 5 — соединительная ресничка; 6 — мешочки палочки
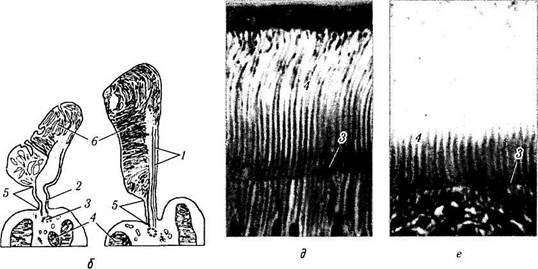
Рис. 5.8.6. Развитие колбочек в эмбриональном периоде и постнатальной жизни (по Albert et al., 1996):
а — 24—26 недель эмбрионального развития; б —плод в возрасте 5 месяцев; в — 45-й месяц постнатальной жизни; г — 13 лет жизни; д, е — взрослые (/ — пигментный эпителий сетчатки; 2 — наружный ядерный слой; 3 —наружная пограничная мембрана; 4 — наружные членики колбочек)
Глава 5. РАЗВИТИЕ ГЛАЗНОГО ЯБЛОКА
 Желтое пятно. Будущая область желтого пятна является центром начала дифференциации сетчатки по направлению от центра к периферии. Именно по этой причине дифференциация нейронов, глиальных клеток и фоторецепторов в этой области обнаруживается довольно рано. Несмотря на это, макулярная область не достигает зрелости в структурном смысле вплоть до 15—45 месяцев после рождения [96].
Желтое пятно. Будущая область желтого пятна является центром начала дифференциации сетчатки по направлению от центра к периферии. Именно по этой причине дифференциация нейронов, глиальных клеток и фоторецепторов в этой области обнаруживается довольно рано. Несмотря на это, макулярная область не достигает зрелости в структурном смысле вплоть до 15—45 месяцев после рождения [96].
К 15-й неделе ганглиозные клетки в области будущей фовеа достаточно дифференцированы и располагаются во внутреннем плексиформном слое. Уже можно увидеть синапсы амакрино-вых, биполярных клеток, а также межклеточные соединения между всеми типами клеток [264]. Приблизительно половина клеток внутреннего ядерного слоя относится к мюллеров-ским и амакриновым клеткам (составляют 35% и 11 % от общего количества клеток). Обнаруживается и небольшая примесь ганглиозных клеток (1% от общего количества). Остальные клетки, видимо, относятся к биполярным клеткам [264]. На этой стадии развития ядра ганглиозных клеток круглые (6,5x8,5 мкм) и окрашиваются бледно в результате диффузного распределения зерен гетерохроматина [264]. Широкий ободок цитоплазмы будущих ганглиозных клеток содержит многочисленные полирибосомы, митохондрии, эндоплазматическую сеть (преобладает шероховатая). Широкие ден-дриты ганглиозных клеток насыщены микротрубочками. Отличием ядер амакриновых клеток является наличие глыбки полового хроматина.
Необходимо отметить, что по мере дифференциации нейронов сетчатой оболочки и образования синаптических соединений между ними возможно формирование специфического нервного импульса различных субпопуляций ганглиозных клеток и других нейронов в ответ на различные раздражители. Показано это в эксперименте на разных позвоночных [81, 156, 166].
На этой стадии развития размер мюллеров-ских клеток в среднем равняется 4,4 х 15,2 мкм [264]. Тела клеток локализуются в средней части внутреннего ядерного слоя. Отростки мюл-леровских клеток интенсивно окрашиваются гистологическими красителями, поскольку они насыщены микрофиламентами и гранулами гликогена.
Фовеа. Область фовеа (рис. 5.8.8, 5.8.9) к 22-й неделе эмбрионального развития представляет собой небольшую зону (приблизительно 1,5 мм в диаметре), содержащую 5—7 рядов клеточных ядер, расположенных в слое ганглиозных клеток. Тонкий слой нервных волокон хорошо отделяет внутренний плексиформный, внутренний ядерный и наружный плексиформный слои от наружного ядерного слоя, содержащего исключительно ядра колбочек [96, 97 139]. Развивающийся слой горизонтальных клеток выглядит в виде полоски бледных круглых
клеток, расположенных снаружи от внутреннего ядерного слоя. Сразу рядом со слоем горизонтальных клеток лежат биполярные клетки. Базофильные ядра мюллеровских клеток занимают середину внутреннего ядерного слоя, а гетерогенное скопление недифференцированных нейронов составляет внутреннюю треть.
В наружном плексиформном слое нежные отростки нейронов вступают в контакт с колбочками, которые к этому моменту времени располагаются в виде одного слоя клеток. Ни внутренние, ни внешние членики еще не дифференцированы, но уже существует наружная пограничная мембрана.
К 24-й неделе эмбрионального развития плотность колбочек в будущей фовеа равна приблизительно 30 000 на мм2. Максимальная плотность палочек (59 200 на мм2) обнаруживается в области, окружающей скопление колбочек [54, 55].
Углубление в центральной области будущей макулы появляется на 24—26-й неделе и связано с истончением слоя ганглиозных клеток и внутреннего ядерного слоя [96, 97, 139]. Количество слоев ганглиозных клеток уменьшается до трех или четырех.


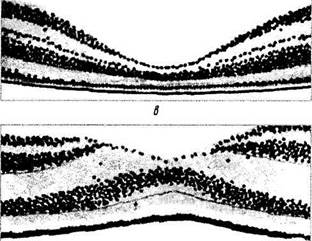
Рис. 5.8.8. Схема развития области желтого пятна сетчатой оболочки (по Seefelder, 1910):
а — 5—6-й месяцы развития; б — 8-й месяц развития; в — ма-кула новорожденного; г — макула взрослого
Развитие сетчатой оболочки

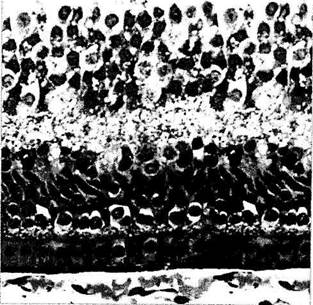
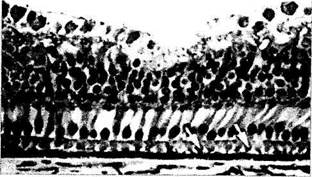
Рис. 5.8.9. Микроскопические особенности развивающейся макулярной области (по Hendrickson Yuodelis 1984):
а — срез через фовеа 22-недельного эмбриона. Слой фоторецепторов содержит исключительно колбочки. Дифференциация колбочек на наружный и внутренний членики на этой стадии отсутствует. Все нейроны и ганглиозные клетки располагаются в наружном слое; б — срез через фовеа на 5-й день после рождения. Слой фоторецепторов содержит колбочки с четкими ядрами. Колбочки обладают большим светлым округлым внутренним члеником, проникшим через наружную пограничную мембрану (стрелки). Наружные членики колбочек прилежат к клеткам пигментного эпителия
На склоне ямки (фовеа) с назальной стороны внутренний ядерный слой не содержит клеток и соответствует слою Чивица остальной части сетчатки.
Проксимальные концы колбочек постепенно суживаются и разворачиваются в боковую сторону. Дифференциация внутренних члеников колбочек сопровождается накоплением в цитоплазме клеток большого количества митохондрий.
К седьмому месяцу внутренний ядерный слой заметно истончается, особенно в области фовеа. Увеличивается и плотность колбочек.
К 8-му месяцу остается только два слоя ганглиозных клеток, а у новорожденных только один.
На момент рождения переходный слой Чивица еще сохранен. Состоит он из аксонов биполярных клеток, направляющихся к внутреннему плексиформному слою. Колбочки вне фо-веолы более зрелые. Даже спустя неделю после рождения толстым колбочкам все еще недостает наружных члеников.
К 4-му месяцу после рождения все слои смещены к периферии относительно склона фовеа. Остаются только ядра колбочек.
В последующем продолжается ремодуляция элементов в области фовеа до тех пор, пока не исчезнет слой Чивица. Происходит это только к 4 годам после рождения [96, 97, 139].
С момента рождения до 4-летнего возраста колбочки удлиняются, а их диаметр уменьшается. Удлинение колбочек коррелирует с развитием наружных члеников и аксонов (волокнистый слой Хенле). Перемещение фоторецепторов в области фовеа приводит к нарастанию плотности колбочек (от 18 на первой неделе после рождения до 42 на 100 мкм у взрослых) [290]. Развитие периферии сетчатки. Граница между эпителием ресничной области и сетчаткой с образованием зубчатой линии (ora serrata) появляется на четвертом месяце эмбрионального развития [6, 14, 49, 139]. К шестому месяцу периферия сетчатки состоит только из тонкого слоя нервных волокон. Зона между зубчатой линией и экватором расширяется вплоть до двухлетнего возраста [135, 142, 173, 201, 249]. Развитие сосудов сетчатки. Особенности эмбриогенеза кровеносных сосудов сетчатой оболочки довольно интенсивно изучались у экспериментальных животных и человека. Основные закономерности этого процесса сводятся к следующему.
Терминальная часть эмбриональной глазной артерии проникает в эмбриональную щель при размере эмбриона, равном 5 мм. Несмотря на то, что эмбриональная щель закрывается между пятой и седьмой неделями эмбрионального развития, терминальная часть эмбриональной глазной артерии остается в зрительном бокале и получает название гиалоидной артерии. Этот сосуд кровоснабжает хрусталик и сетчатку.
К четвертому месяцу вокруг гиалоидной артерии скапливаются мезенхимные клетки, часть которых направляется к диску зрительного нерва. Ранее эти клетки проникают во внутренние слои сетчатки в виде плотных тяжей. По ходу распространения тяжей образуются сосуды [6, 14, 49, 139, 148]. Появляются межклеточные контакты между эндотелиальными клетками. Уже на ранних стадиях развития можно обнаружить также перициты.
Развивающийся сосуд окружен базальной мембраной, с которой контактируют отростки астроцитов [70]. Особенностью развивающейся
Глава 5. РАЗВИТИЕ ГЛАЗНОГО ЯБЛОКА
сосудистой сети является наличие большого количества веретеновидных клеток, являющихся предшественниками структурных элементов сосудов, и фагоцитирующих клеток, играющих роль в ангиогенезе [67, 69].
Процесс васкуляризации сетчатки распространяется от ее центральных участков к периферии [106]. Уже к пятому месяцу сосуды распространяются до верхней и нижней височной половины сетчатки. С назальной стороны развитие сосудов задерживается (рис. 5.8.10).
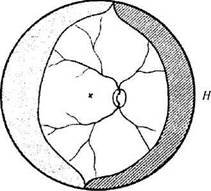
Рис. 5.8.10. Распределение сосудов сетчатки в плоскости сетчатой оболочки к 5-му месяцу эмбрионального развития (объяснение в тексте)
В области склона фовеа появляются маленькие сосуды в слое ганглиозных клеток между 24-й и 26-й неделями эмбрионального развития. Затем формируется примитивная капиллярная сеть, состоящая из артериол и вен. Капилляры достигают зубчатой линии к восьмому месяцу, а полное вызревание сосудистой сети отмечается лишь к 3-му месяцу после рождения.
Начало образования сосудов сетчатки проявляется перемещением от диска зрительного нерва многочисленных веретенообразных ме-зенхимных клеток, которые и являются предшественниками эндотелиальных клеток сосудов. В последующем они образуют тяжи и появляется просвет сосуда. Процесс древовидного ветвления формирующихся сосудов первоначально происходит во внутренних слоях сетчатки и вблизи диска зрительного нерва. Затем сосуды распространяются к периферии сетчатки и височно. «Созревание» сосудистого сплетения характеризуется существенным преобразованием сосудов, сопровождающимся уменьшением количества незрелых эндотелиальных клеток. Иной тип формирования сосудов обнаруживается в наружных слоях сетчатки. Возникают они путем «вытяжения» зародышевых капиллярных сосудов уже от существующего внутреннего слоя сосудов. Этот процесс начинается вблизи фовеа [106].
Нами указывалось на наличие в процессе развития сетчатки достаточно большого количества перицитов, расположенных вокруг фор-
мирующихся сосудов. Первые перициты можно обнаружить уже на пятом месяце эмбриогенеза, но наибольшее их количество выявляется только в постнатальном периоде. Происхож; дение этих клеток до конца не ясно. Эти клетки могут являться видоизмененными клетками эндотелия, сохраненными после ретракции эмбриональной сосудистой сети. Однако их присутствие на ранних этапах эмбриогенеза подтверждает гипотезу их происхож
|
|
|
