 |


Регуляция численности популяций в биоценозах
|
|
|
|
8.6.1. Модификация и регуляция популяций
Современная теория рассматривает динамику численности популяций как авторегулируемый процесс. Любой популяции организмов в конкретных условиях свойствен определенный средний уровень численности, вокруг которого происходят колебания. Отклонения от этого среднего уровня имеют разный размах, но в норме после каждого отклонения численность популяции начинает изменяться с обратным знаком. Теория динамики численности разрабатывается в основном на примере животных, хотя целый ряд ее положений применим и к растениям, грибам, простейшим и прокариотам.
Выделяют две принципиально разные стороны популяционной динамики: модификацию и регуляцию. Модификация – это случайное отклонение численности, возникающее в результате воздействия самых разнообразных факторов, не связанных с плотностью популяции. Регуляция – это возврат популяции после отклонения к исходному состоянию, совершающийся под влиянием факторов, сила действия которых определяется плотностью популяции.
Модифицирующие факторы, вызывая изменение численности популяций, сами не испытывают влияния этих изменений. Действие их, таким образом, одностороннее. К ним относятся все абиотические влияния среды на организмы, на качество и количество их корма и т. п. Благоприятная погодная обстановка может послужить причиной массовой вспышки размножения вида и перенаселения занимаемой им территории, как, например, в случае стадных саранчовых. Отрицательное воздействие модифицирующих факторов, наоборот, снижает численность популяции иногда до полного ее исчезновения.
Климатические и погодные изменения оказывают и прямое, и опосредованное влияние на живые организмы. На популяциях это влияние проявляется через усиление или ослабление смертности. Так, в нераспаханных степях большим бедствием для копытных был «джут» – сочетание многоснежья с гололедицей. В такие зимы наблюдался массовый падеж скота, сайгаков, дроф, стрепетов, не способных добывать корм из-под снега. В малоснежные холодные зимы в лесной зоне из-за промерзания грунта в массе гибнут кроты, полевки, мыши. Заморозки в начале лета могут повлиять на выживаемость птиц. У тетеревов и глухарей, например, замерзают в гнездах яйца, когда самки отлучаются для кормежки. Насекомоядные птицы в это время гибнут от голода. Засухи и наводнения губят многих насекомых, их личинок и кладки. В выжженной солнцем степи после миграции по ней сайгаков остается множество трупов этих животных, преимущественно молодняка.
|
|
|
Влияние модифицирующих факторов, не зависящих от плотности популяции, может вызывать резкий спад ее численности. В сельском хозяйстве чрезвычайно важно прогнозирование погодных условий для возможностей защиты урожая.
Численность многих видов проявляет прямую корреляцию с ходом температурных условий, что особенно наглядно для пойкилотермных. Из-за выраженного сильного и разнонаправленного действия на популяции погодных условий ряд экологов поддерживал в прошлом так называемую «климатическую» теорию динамики численности. По этой теории именно вариации погодных факторов определяют пределы колебания численности видов. Однако дальнейшие исследования показали, что никакие комбинации случайных, односторонне действующих факторов не могут удерживать популяции в тех определенных границах, которые характерны для них в конкретных биоценозах. Факторы, действие которых проявляется на уровне организмов и не зависит от плотности популяций, не могут быть регуляторами их численности.
|
|
|
Регулирующие факторы не просто изменяют численность популяции, а сглаживают ее колебания, приводя после очередного отклонения от оптимума к прежнему уровню. Это происходит потому, что эффект их воздействия тем сильнее, чем выше плотность популяции. В качестве регулирующих сил выступают межвидовые и внутривидовые отношения организмов.
Межвидовые связи осуществляются в биоценозах, поэтому относятся к группе биоценотических механизмов регуляции численности популяции. Наиболее эффективные из них – трофические отношения организмов: хищничество, паразитизм, собирательство, пастьба и другие, как прямые, так и косвенные. Прямые связи хищник – жертва – наиболее изученные регуляторные механизмы в сообществах.
Общее число жертв, которых может уничтожить один хищник, сначала растет почти пропорционально росту численности потребляемого вида. Это так называемая функциональная реакция хищников на жертвы. Однако она имеет предел, обусловленный физическими возможностями потребителя (рис. 135). После полного насыщения хищники обычно слабо реагируют на жертву. Новые возможности влиять на популяции жертв возникают при росте численности хищников за счет успешного размножения на хорошей кормовой базе. Это явление получило название численной или количественной реакции хищников на рост численности жертв. Количественная реакция хищников всегда запаздывает по сравнению с ходом размножения жертвы, так как является ответом на изменение кормовой базы.

Рис. 135. Функциональная реакция одиночной самки хальциды Dahlbomi-nus, заражающей коконы пилильщика Neodiprion sertifer (по Дж. Варли и др., 1978): слева – число коконов, пораженных одной самкой паразита, при разной их плотности; справа – зависимость процента пораженных коконов от их плотности при постоянной численности паразита
В лабораторных условиях при совместном содержании хищника и его жертвы отмечаются периодические изменения численности обоих видов, имеющие довольно закономерный характер. В опытах, проведенных нашим отечественным биологом Г. Ф. Гаузе в 30-х годах с пробирочными культурами простейших, было обнаружено следующее. Усиленное размножение хищной инфузории Didinium nasutum закономерно следовало всякий раз за размножением ее жертвы – туфельки Paramecium caudatum. Когда хищник достигал высокой численности, он полностью уничтожал парамеций, после чего наступала гибель самих Didinium, лишенных пищевой базы. Однако, когда в пробирки помещали некоторое количество песчинок, под которыми часть туфелек могла найти недоступные для хищников убежища, после гибели Didinium наступало новое массовое размножение парамеций (рис. 136). В другой серии опытов, когда культивировали разные виды парамеций, а в качестве их пищи – дрожжевые клетки рода Sacharomyces, было получено при дополнительных условиях несколько закономерных циклов колебаний численности дрожжей и туфелек. Механизмы подобных колебаний можно представить следующим образом.
|
|
|
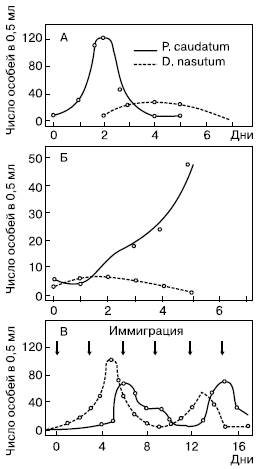
Рис. 136. Изменение численности инфузорий Paramaecium caudatum (жертва) и Didinium nasutum (хищник) при культивировании в общем сосуде (по Г. Ф. Гаузе, 1934): А – нет убежищ для парамеций; Б – парамеции имеют убежища от хищников; В – с повторным добавлением обоих видов, или иммиграцией
Увеличение численности жертв происходит до тех пор, пока темп выедания их нарастающим количеством хищников превысит скорости размножения. После этого дальнейшая деятельность хищников только подрывает кормовую базу, что ведет к снижению их собственной численности. Это позволяет немногим оставшимся в живых жертвам вновь размножиться и, следовательно, улучшить кормовые условия для хищников. Если последние еще присутствуют в системе, новое увеличение количества жертв способствует новому подъему численности хищников и цикл повторяется снова.
В природных условиях такие закономерные циклические изменения численности двух видов, связанных пищевыми отношениями, выявить трудно, так как на их размножение и гибель влияют взаимодействия со множеством других видов, а также абиотические изменения среды. Однако в ряде случаев и в природной обстановке удается заметить регулярные периодические изменения обилия хищников и их жертв.
Особенно наглядно такие колебания обнаруживаются в обедненных сообществах, где межвидовые связи не столь разнообразны: в тундрах и полярных пустынях, лесах с господством одной породы деревьев, культурных садах и т. п. Циклические изменения численности выявлены, например, у леммингов и их основных потребителей – песцов, массово размножающихся хвое– и листогрызущих насекомых и их паразитов, вредителей яблонь – паутинных клещиков и питающихся ими хищных клещей и у многих других сопряженных пищевыми связями видов. Более детальные исследования показывают, что циклические колебания численности жертв чаще связаны с проявлением внутрипопуляционных механизмов гомеостаза, но и взаимоотношения с хищниками вносят свой вклад в размах этих колебаний.
|
|
|
Таким образом, истребление жертв и поражаемость хозяев зависят от численности врагов, но и обилие последних также определяется численностью объектов их питания.
Паразиты – также мощный фактор сдерживания численности хозяев. Они, как правило, редко вызывают массовую гибель пораженных особей. Но поскольку зараженность паразитами ослабляет организм хозяина, она приводит, как правило, к падению плодовитости или даже полной стерилизации, что имеет следствием снижение численности следующих поколений.
Некоторые паразиты, однако, обязательно приводят хозяина к гибели, что сокращает численность заражаемого ими поколения. Эту группу паразитов называют часто паразитоидами. К ним относятся, например, насекомые, откладывающие яйца в яйца и личинок других насекомых.
Возбудители инфекционных заболеваний также сильно зависят от плотности популяций поражаемых ими видов, так как при достижении последними определенного порога численности становится возможной легкая передача инвазионного начала. Поэтому инфекционные болезни (тоже основанные на трофических связях) распространяются, как правило, внезапно и взрывообразно.
Конкуренты также являются регуляторами плотности популяций в биоценозах, так как взаимно обусловливают численность друг друга, вынуждая ограничивать масштабы использования ресурсов.
Биоценотические механизмы являются внешними по отношению к популяции регуляторами, сдерживающими рост ее численности.
Внутренние механизмы – это все те качественные изменения популяции в ответ на повышение плотности, которые рассматривались как механизмы ее гомеостаза. Многообразие этих механизмов (самоизреживание, каннибализм, территориальное поведение, расселительные инстинкты, фазовость насекомых, стресс-реакция млекопитающих, изменения плодовитости, агрессивности и т. п.) свидетельствует о том, что любой способ снятия угрозы перенаселения поддерживается естественным отбором как важная адаптация в жизни вида.
|
|
|
8.6.2. Инерционная и безынерционная регуляция
Разные типы отношений определяют быстроту ответных реакций на изменения численности популяций. В связи с этим среди факторов популяционной динамики выделяют инерционные и безынерционные регуляторные механизмы.
Инерционные механизмы зависят главным образом от плотности предыдущих поколений, безынерционные – от плотности текущих генераций. Например, функциональная реакция хищников – безынерционный механизм воздействия на популяцию жертв, так как количество пойманных жертв увеличивается сразу вслед за ростом их численности. Количественная же реакция, связанная с увеличением численности самих хищников, всегда запаздывает, так как для миграции или размножения их нужно обычно значительное время. Часто поэтому естественные враги сильно размножаются уже тогда, когда численность жертв по каким-то причинам пошла на убыль. В динамике численности насекомых, например, наибольшую инерционность имеет деятельность энтомофагов, особенно тех, у кого время генерации больше, чем у жертвы. Менее инерционны болезнетворные микроорганизмы. При высокой плотности популяций насекомых эпизоотии могут «вспыхивать» очень быстро. При этом играет роль и скученность насекомых, и их ослабленность в результате начинающегося недостатка корма.
Внутрипопуляционные регуляторные механизмы тоже отличаются по степени инерционности. Конкуренция, каннибализм, миграции достигают эффекта в одном поколении и мало зависят от плотности предыдущих. Такие же явления, как диапауза, изменение возрастного состава и соотношения полов, усиление полиморфизма, сказываются на численности не текущего, а следующих поколений. Здесь также проявляется эффект запаздывания. К этой же группе относятся механизмы сигнального действия роста плотности популяции, которое влечет за собой изменения физиологического состояния и поведения животных еще до проявления острых конкурентных отношений, такие, как фазовость насекомых, стресс у млекопитающих и др.

Рис. 137. Пороги и зоны активности основных механизмов регуляции численности насекомых (по Г. А. Викторову, 1976)
Ограничение возможных колебаний численности популяций имеет большое значение не только для их собственного процветания, но и для устойчивого существования сообществ. Успешное сожительство организмов разных видов возможно только при их определенных количественных отношениях. Естественным отбором закреплены поэтому самые разнообразные заслоны на пути катастрофического увеличения численности популяций, регуляторные механизмы имеют множественный характер. Г. А. Викторов дал схему для иллюстрации действия регулирующих факторов на насекомых (рис. 137). Если темпы роста популяции не очень высоки, то для сдерживания ее численности достаточно деятельности многоядных (т. е. неспециализированных) хищников, у которых данный вид составляет лишь часть рациона. При более быстром увеличении численности насекомых хищники, не успевая выедать весь прирост, теряют регулирующую роль. Однако высокая плотность популяции благоприятна для массового размножения специализированных паразитов, которым становится легче находить хозяев. При еще более быстром темпе роста популяции паразиты также теряют регулирующую роль, но повышается вероятность вспышки инфекционных заболеваний, так как возбудители болезней быстро распространяются при частых контактах особей. Наряду с этим приходят в действие внутрипопуляционные способы поддержания гомеостаза, различные у разных видов. Предельно возможный рост популяции – до полного исчерпывания ресурсов среды и подрыва своего дальнейшего существования. В природных популяциях такие катастрофические события не происходят из-за множественности регулирующих воздействий межвидового и внутривидового характера, включающихся в разных интервалах плотности. Сложная система связей как бы страхует популяцию от превышения границ оптимальной численности в данном сообществе. В технике системы с множественным обеспечением называют ультрастабильными. Регуляция их осуществляется по принципу отрицательной обратной связи: отклонения от исходного режима работы вызывают в системе такие изменения, которые вновь возвращают ее к исходному состоянию.
Динамика численности популяций в естественной обстановке – это также автоматически регулируемый процесс, механизмы стабилизации которого отработаны длительной историей совместного развития видов (рис. 138). В сообществах, искусственно создаваемых человеком или упрощенных в результате антропогенных воздействий, регуляторные связи ослаблены, и поэтому в них возможны катастрофические для биоценоза размножения отдельных видов – вредителей сельскохозяйственных растений и лесных насаждений, грызунов, паразитов, возбудителей болезней и т. п.

Рис. 138. Общая схема динамики численности насекомых (по Г. А. Викторову, 1976)
Природная регуляция численности имеет две особенности. Во-первых, большинство регуляторных механизмов действует в ответ на уже происшедшее изменение численности популяции и регуляторный эффект достигается с некоторым замедлением. Это значит, что полной стабилизации достигнуто быть не может, численность популяции всегда совершает колебания, синхронные с воздействием модифицирующих факторов, а регулирующие лишь уменьшают размах возможных колебаний. Таким образом, популяциям свойствен тип регуляции, управляемой ошибкой: механизмы регуляции срабатывают лишь при выходе численности за определенные пределы.
Во-вторых, регуляция способствует преимущественно лишь активному ограничению роста популяции, тогда как подъем численности после сильного снижения возможен в результате уменьшения силы действия регуляторов.
Разграничение факторов динамики численности организмов на модифицирующие и регулирующие имеет большое практическое значение из-за принципиальных различий их влияния на популяцию.
Изучение модифицирующих факторов важно для выяснения причин колебания численности и их прогнозирования, изучение регуляторных механизмов – для ограничения амплитуды колебания численности и ее максимально возможной стабилизации.
Таким образом, как масштабы, так и ход колебания численности любого вида в природных сообществах исторически обусловлены естественным отбором в зависимости от особенностей биологии, характера внутривидовых связей и межвидовых отношений.
Как правило, численность видов, имеющих много врагов и конкурентов, ограничивается в биоценозах именно внешними регуляторами, их функциональными и количественными реакциями. Собственные механизмы гомеостаза выявляются лишь при очень высокой плотности популяций, когда возникают сбои в активности потребителей. Виды, которых «все едят», отзываются лишь на чрезмерное увеличение собственной численности, грозящей подрывом ресурсов. Наоборот, крупные хищники и другие виды, испытывающие слабый пресс естественных врагов, обладают очень эффективными внутривидовыми способами регуляции плотности популяции.
8.6.3. Типы динамики численности популяций
Можно выделить три основных типа популяционной динамики численности (рис. 139).
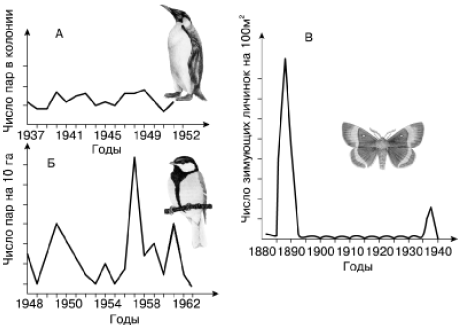
Рис. 139. Основные типы динамики численности популяций (по М. Уильямсону, 1975): А – пингвина великолепного; Б – большой синицы; В – зимующих гусениц соснового шелкопряда
I. Стабильный тип – отличается небольшим размахом колебаний (в несколько раз, однако не на несколько порядков величин). Свойствен видам с хорошо выраженными механизмами популяционного гомеостаза, высокой выживаемостью, низкой плодовитостью, большой продолжительностью жизни, сложной возрастной структурой, развитой заботой о потомстве. Целый комплекс эффективно работающих регуляторных механизмов держит такие популяции в определенных пределах плотности. Такова, например, динамика численности крупных млекопитающих и птиц, а также ряда беспозвоночных.
II. Флюктуирующий тип – колебания происходят в значительном интервале плотностей, различающихся на один-два порядка величин. При этом различают три фазы колебательного цикла: нарастания, максимума, разрежения численности. Возврат к стабильному состоянию происходит быстро. Регуляторные механизмы не теряют контроля за численностью популяций, увеличивая свою эффективность вслед за увеличением плотности. Преобладают слабоинерционные меж– и внутривидовые взаимодействия. Такой ход численности широко распространен в разных группах животных.
III. Взрывной тип с вспышками массового размножения – прекращение действия модифицирующих факторов не вызывает быстрого возврата популяции в стабильное состояние. Динамика численности складывается из циклов, в которых различают пять обязательных фаз: нарастания численности, максимума, разреживания, депрессии, восстановления. Для популяций периодически характерны предельно высокий и необычайно низкий уровень численности. По фазам цикла также сильно меняются показатели размножения, возрастной и половой структуры популяции, физиологического состояния, поведения, а иногда и морфологических особенностей составляющих ее особей. Такой ход численности обнаруживается чаще всего у видов с малой продолжительностью жизни, высокой плодовитостью, быстрым оборотом генераций. Он свойствен, например, некоторым насекомым (саранчовые, вредители леса – усачи, короеды, ряд чешуекрылых и пилильщиков и др.), среди млекопитающих отмечен у многих видов мышевидных грызунов.
Тип динамики численности – скорее популяционная, но не видовая характеристика. Популяции одних и тех же видов в различных условиях могут характеризоваться разным ходом динамики численности. Это объясняется преимущественно тем, что среди регуляторных механизмов большую роль играют межвидовые взаимосвязи, которые в пределах ареала вида могут быть разной степени напряженности. Так, многие виды, которые в природных условиях сдерживались врагами, проявляют склонность к вспышкам массового размножения в садах и на полях, где ослаблен биологический контроль.
8.6.4. Механизмы динамики численности
Типы популяционной динамики численности наиболее хорошо изучены у лесных насекомых, на примере которых выявляется большое многообразие конкретных связей и особенностей видов, влияющих на плотность их популяций.
В лесных сообществах преобладают виды со стабильным ходом популяционной динамики. Они питаются практически за счет всех частей деревьев (стволы, корни, побеги, листья, почки, плоды, семена), однако при этом не оказывают губительного воздействия на растения. Как правило, такие индифферентные виды отличаются низкой миграционной активностью, ведут оседлый образ жизни. Комплекс их паразитов включает обычно несколько сильно специализированных видов, эффективно поражающих популяцию хозяина.
Флюктуирующий тип динамики численности свойствен многим ксилофагам (потребителям коры и древесины): усачам, златкам, короедам. Для них характерно совместное заселение кормовых объектов – ослабление деревьев. Это позволяет быстро снизить устойчивость дерева, но в то же время совместное обитание ксилофагов обостряет конкурентные отношения между ними, что действует как безынерционный механизм регуляции численности.
Для короедов, поселяющихся первыми на ослабленных деревьях, при избыточном запасе питательных веществ (растворимых углеводов и крахмала) развитие лимитируется воздействием защитных реакций дерева, таких, например, как смоловыделение у хвойных. Кроме того, еще отсутствуют симбиотические микроорганизмы. Сопротивление еще жизнеспособных деревьев может быть сломлено при концентрированном нападении и избыточно высокой плотности вредителя. Другой предел пригодности дерева для короедов – полное отмирание и разрушение луба. Между двумя указанными состояниями дерева для короедов создается оптимум кормовых условий, отмечается максимальная выживаемость всех фаз развития и наиболее высокие коэффициенты размножения.
Относительную роль различных регуляторных механизмов, определяющих флюктуирующий ход численности лесных насекомых, можно проследить на примере большого лиственного короеда – Ips subelongatus. Этот вредитель лиственницы и частично других пород широко распространен по всей таежной зоне.
Жуки нападают на физиологически ослабленные живые лиственницы, у которых отсутствует активное защитное смоловыделение. Внедряясь под кору, самцы продуцируют запаховые вещества – феромоны, привлекающие других особей, чем обеспечивается массовость заселения подходящего дерева и его дальнейшее ослабление. Феромоны играют также роль химических регуляторов внутрипопуляционных отношений – распределения короедов по стволу, своего рода меткой территории отдельных гнезд. Жуки, не нашедшие места на данном дереве, переселяются на другие. В дальнейшем внутривидовая конкуренция регулируется самками, которые в зависимости от плотности поселения изменяют длину маточных ходов и количество откладываемых яиц. При возрастающем недостатке площади часть самцов и самок мигрирует на другие объекты. Личинка короеда за период развития потребляет 0,78 см6 луба. Если «кормовые площадки» личинок при высокой плотности поселения становятся меньше этого предела, среди них увеличивается смертность. Таким образом, внутривидовая конкуренция регулирует заселенность дерева на всех фазах жизни вредителя: на стадии взрослых жуков она ведет к усилению миграций, на стадии личинок – к гибели части популяции. Показатели размножения остаются высокими на всех этапах флюктуации численности популяции.
Естественных врагов данного вредителя можно разделить на пять групп, по особенностям их взаимодействия с короедом. 1. Враги, действующие в период лёта короедов, – птицы (синицы, дятлы и др.), ктыри, муравьи. Они уничтожают часть популяции, но одновременно и энтомофагов короеда, т. е. других его естественных врагов. Их регуляторная деятельность поэтому слабо эффективна. 2. Хищные насекомые, уничтожающие короеда на всех фазах развития под корой (ряд пестрянок, чернотелок, карапузиков, хищных мух и клопов). При низкой и средней плотности вредителя хищники сдерживают рост его численности, при высокой же плотности короедов – способствуют повышению выживаемости их вследствие ослабления внутривидовой конкуренции. 3. Паразиты, заражающие только личинок короедов, – бракониды, хальциды. На регуляцию численности воздействуют слабо, так как пораженные личинки погибают лишь в конце жизни и общая смертность их от паразитов составляет несколько процентов. 4. Паразиты, заражающие взрослых жуков, – нематоды, некоторые перепончатокрылые. Их влияние проявляется в снижении плодовитости и сокращении продолжительности жизни жуков, регулирующий эффект в целом невысок. 5. Бактериальные и грибные заболевания. Эпизоотии среди короедов возникают лишь при резком ухудшении условий, когда действуют более мощные механизмы снижения численности.
Таким образом, в межвидовых отношениях короедов более или менее значительную роль в регуляции их численности имеют лишь хищные насекомые при относительно невысокой численности вредителя.
Способность к вспышкам массового размножения свойственна относительно немногим видам лесных насекомых. В сибирской тайге у скрытоживущих видов взрывной тип динамики численности характерен для алтайского лиственничного усача, большого черного хвойного усача, лиственничной почковой галлицы и некоторых других. Среди открытоживущих листогрызущих насекомых способность давать вспышки массового размножения свойственна лишь некоторым видам чешуекрылых и перепончатокрылых (пилильщикам, ткачам). Отличительные черты экологии таких видов: высокая выживаемость в сильно изменчивой среде благодаря специальным адаптациям, высокой миграционной активности, высокой и изменчивой плодовитости. У открытоживущих видов часто выявляется эффект группы и фазовая изменчивость.
Один из опаснейших вредителей хвойных пород – сибирский шелкопряд Dendrolimus sibiricus, распространенный от Урала до Тихого океана. Этот вид обладает следующими внутрипопуляционными механизмами регуляции численности.
В очагах сибирского шелкопряда по фазам вспышки существенно изменяется соотношение полов. Доля самок варьирует от 32 до 76 %. При нарастании вспышки доминируют самки, при затухании – самцы. В переуплотненных популяциях повышается смертность самок на всех фазах развития, а также отмечается их более высокая миграционная активность из очагов размножения. В фазе максимума численности на периферии очага доля самок составляет до 73 %, а в центре – 44 %.
У сибирского шелкопряда, как и у других видов с вспышками массового размножения, плодовитость изменяется в широких пределах. Этот показатель сильно зависит от плотности популяции (рис. 140). При низкой и сверхвысокой численности плодовитость самок снижается почти втрое. Наиболее высокая плодовитость наблюдается в фазе нарастания численности (свыше 300 яиц на самку), а затем закономерно снижается под влиянием конкуренции и недостатка корма в переуплотненных очагах. Изменение соотношения полов и плодовитости можно рассматривать как безынерционные или слабоинерционные механизмы регуляции численности. Их действие проявляется в течение жизни одного поколения.

Рис. 140. Фазовый портрет динамики численности сибирского шелкопряда в Красноярском крае (из А. С. Исаева и др., 2001):
по вертикали – логарифм коэффициента размножения;
по горизонтали – логарифм плотности популяции гусениц на одно дерево. Схема иллюстрирует цикличность вспышек и зависимость показателей размножения от плотности популяции
Миграционная активность сибирского шелкопряда – также эффективный внутрипопуляционный механизм регуляции численности. У этого вида отмечены разные типы массовых перемещений. В период низкой численности пульсирующие миграции перераспределяют вредителя на площади лесных массивов, в результате чего формируются резервации – места сохранения вида. В основном это перестойные пихтовые леса на плакорах или склонах южной экспозиции. В начальной фазе вспышки происходит стягивание вредителя с больших территорий на ограниченный участок леса с наиболее благоприятными условиями (конденсирующие миграции). К концу фазы нарастания численности начинается вылет бабочек из первичных очагов, где к этому времени плотность популяции достигает критических величин. На фазе максимума численности происходят массовые выселения бабочек и их перемещение воздушными потоками на расстояния до 100 км. Среди мигрантов преобладают самки с запасом зрелых яиц. В редких случаях отмечаются активные массовые миграции бабочек. При затухании очагов сибирского шелкопряда развиваются дисперсионные миграции – рассеивание популяции из очагов размножения в окружающие лесные массивы.
Для сибирского шелкопряда известен также эффект группы – положительное влияние оптимальной плотности, которое проявляется в основном у гусениц I–III возрастов и выражается в ускорении их развития более чем в 2 раза. В регуляцию хода численности вредителя вносит вклад также гетероцикличность – наличие особей с одно– и двухлетним циклом в зависимости от плотности популяции.
Повышение плотности очага сильно ускоряет метаболические процессы у членов популяции, что выражается как в ускорении сроков развития, так и в изменении поведения и внешнего вида бабочек и гусениц – появляются особи с темной окраской. В разных фазах цикла массового размножения количество темных бабочек в лесах Средней Сибири изменяется от 10 до 80 %.
Среди естественных врагов сибирского шелкопряда отмечено более 60 видов. Однако, несмотря на разнообразие, энтомофаги не могут постоянно держать под контролем численность вредителя из-за высокой инерционности их количественной реакции. Усложняющий фактор – высокая активность сверхпаразитов. Многие энтомофаги сибирского шелкопряда гораздо менее его устойчивы к факторам абиотической среды. Например, один из эффективных паразитов яйцеед-теленомус в массе гибнет в холодные зимы. Вот почему вспышки размножения сибирского шелкопряда часто возникают после холодных малоснежных зим.
При низкой численности сибирского шелкопряда в резервациях за счет естественных врагов-энтомофагов гибнет до 95 % яиц, 40 % гусениц и 70 % куколок. Основными регуляторами при этом являются яйцеед-теленомус и тахина-мазицера. Общие показатели смертности в популяциях достигают более 99 %. Это свидетельствует о высоких возможностях вредителя переходить к вспышке. Быстрый рост численности может быть обеспечен даже небольшим (5-10 %) увеличением выживаемости. На фазе нарастания численности поражаемость от энтомофагов падает (яйца – до 50 %, гусеницы – до 11 %, куколки – до 23 %). Энтомофаги «не успевают» сдерживать растущую популяцию вредителя. В период кульминации вспышки пресс энтомофагов на популяцию шелкопряда усиливается и достигает наибольшего выражения в период разреживания и депрессии численности, когда естественные враги полностью уничтожают избыток особей.
Таким образом, при массовых вспышках размножения лесные насекомые-вредители выходят из-под межвидового контроля и их возврат к стабильной численности происходит в основном в результате внутрипопуляционных регулирующих механизмов, которые наиболее эффективно действуют лишь при очень высоких уровнях плотности, губительной для лесного насаждения.
Понимание и учет всех этих факторов, определяющих динамику численности лесных вредителей, совершенно необходимы для принятия действенной системы мер борьбы с ними и профилактики вспышек, для определения необходимости, сроков, доз и форм применения средств, сдерживающих массовое размножение. С экологической точки зрения задача заключается в удержании популяции в зоне депрессии численности.
Понимание механизмов, определяющих движение численности популяций, дает в руки ключ управления ими. Однако эта задача не может решаться трафаретно для разных видов. Во всех случаях необходимо детальное знание экологических особенностей конкретного вида и условий его биотического окружения, а также характер взаимодействия модифицирующих факторов среды.
8.6.5. Влияние ослабления или усиления пресса хищников на динамику популяций и структуру сообществ
Ярким примером того, к чему приводит снятие пресса потребителей на популяции жертв, являются масштабные «нашествия» чужеродных видов, попадающих в подходящие для размножения условия на других материках. На родине такие виды испытывают влияние активных регуляторов, занимают соответственные экологические ниши в биоценозах и, как правило, не считаются вредителями в хозяйственной практике человека. В новой среде, не имея специализированных врагов, эти виды за короткие сроки дают взрывы численности, нарушая структуру сложившихся сообществ. С экологических позиций впервые обзор и анализ таких событий был сделан английским зоологом Ч. Элтоном, одним из основоположников популяционной экологии. Вводя понятие «взрыв» применительно к непомерному увеличению численности какого-либо вида, Ч. Элтон объяснял, что он намеренно употребляет это слово, так как оно обозначает «внезапное освобождение сил, проявление которых сдерживалось ранее другими силами».
«Нашествия» иноземных видов, занос которых преднамеренно или случайно осуществляется человеком, широко распространены в современном мире и часто вызывают большие осложнения в хозяйственной деятельности. Относительно немногие примеры успешной борьбы с ними связаны, как правило, с интродукцией естественных врагов, т. е. воссозданием системы биоценотической регуляции. Если на «захваченной» видом территории постепенно складывается новый комплекс регуляторов, тогда его численность идет на убыль. Однако для некоторых вселенцев этот процесс происходит достаточно медленно. Таким примером является опасный вредитель картофеля колорадский жук Leptinotarsa septemlineata, распространение которого в Европе началось после заноса из Америки в 20-х годах прошлого века с западных берегов Франции. Постепенно распространяясь на восток, колорадский жук занял в настоящее время всю европейскую территорию, картофелеводческие районы Сибири, обнаружен на Дальнем Востоке и острове Сахалин (рис. 141). Жук и его личинки имеют ядовитую гемолимфу, спасающую их от насекомоядных птиц и других позвоночных. У колорадского жука на евроазиатской территории до сих пор не сформировался эффективный комплекс потребителей. Потери урожая картофеля от колорадского жука составляют в среднем до 40 %.
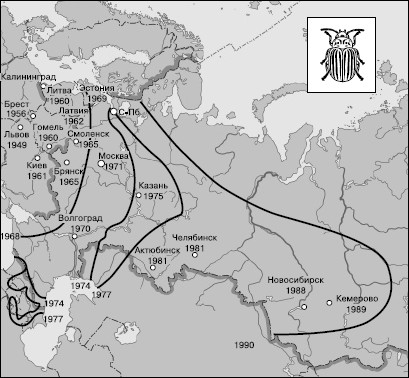
Рис. 141. Распространение колорадского жука по территории России (на 1990 г.)
Другим примером современного «нашествия» вида, изменившего всю структуру сообщества, является размножение в Черном море гребневика Mnemiopsis leigyi, занесенного с балластными водами судов от побережья Северной Америки (рис. 142). Этот гребнев
|
|
|
