 |


Периоды и возрастные состояния в жизненном цикле растений
|
|
|
|
(по Н. М. Черновой, А. М. Быловой, 1988)
| Периоды | Возрастные состояния особей | Принятое обозначение |
| 1. Первичного покоя (латентный) | Покоящиеся семена | |
| II. Предгенеративный (виргинальный) | Проростки (всходы) Ювенильные Имматурные Виргинальные (молодые вегетативные, взрослые вегетативные) | p j im v |
| III. Генеративный | Молодые генеративные Средневозрастные генеративные Старые генеративные | g1 g2 g3 |
| IV.Постгенеративный (старческий, сенильный) | Субсенильные (старые вегетативные) Сенильные | ss s |
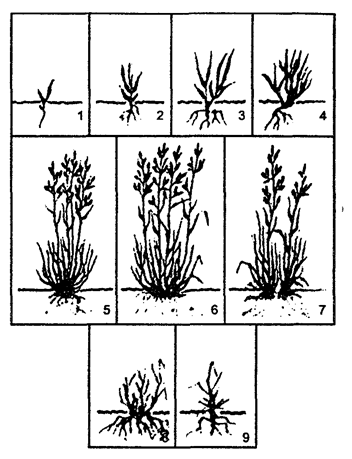
Рис. 9.4. Возрастные группы овсяницы луговой:
1 — проросток; 2, 3, 4 — молодые растения;
5, 6, 7 — взрослые растения, 8, 9 — старые растения
К периоду первичного покоя относятся покоящиеся семена; к предгенеративному — проростки (всходы), ювенильные, имматурные, виргинальные; к генеративному — молодые генеративные, средневозрастные генеративные, старые генеративные; к постгенеративному — субсенильные (старые вегетативные), сенильные.
Проростки имеют смешанное питание за счет как запасных веществ, так и собственной ассимиляции. Для них характерно наличие зародышевых структур: семядолей, зародышевого корня, побега. Ювенильные растения переходят к самостоятельному питанию. У них, например бобовых, уже отсутствуют семядоли, но организация еще проста: листья иной формы и размера, чем у взрослых. Имматурные имеют признаки и свойства, переходные от ювенильных растений к взрослым, происходит смена типов нарастания, начало ветвления и т. д. У взрослых вегетативных растений появляются черты типичной для вида жизненной формы в структуре подземных органов. Листья взрослые, генеративные органы отсутствуют. Молодые генеративные растения развивают генеративные органы, происходит окончательное формообразование взрослых структур. Средневозрастные генеративные растения отличаются максимальным ежегодным приростом и семенной продуктивностью. Старые генеративные растения характеризуются резким снижением генеративной функции, ослаблением процессов корне- и побегообразования. Процессы отмирания преобладают над процессами новообразования. Старые вегетативные растения отличаются прекращением плодоношения. У них возможно упрощение жизненной формы, появление листьев имматурного типа. Сенильные растения крайне дряхлы, при возобновлении реализуются немногие почки, вторично появляются некоторые ювенильные черты: форма листьев, характер побегов.
|
|
|
У растений-монокарпиков, включая однолетних, а иногда и у поликарпиков отсутствует постгенеративный период. Распределение особей ценопопуляции по возрастным состояниям называется ее возрастным спектром. Счетной единицей могут являться отдельные особи, парциальные кусты (длиннокорневые растения) или одиночные побеги (некоторые длиннокорневищные и корнеотпрысковые растения).
Ценопопуляцию, в возрастном спектре которой в момент наблюдения представлены только семена и молодые особи, называют инвазионной. Обычно это молодая ценопопуляция, только что внедрившаяся в фитоценоз. Поддержание ее обеспечивается заносом зачатков извне. Ценопопуляция называется нормальной, если она представлена всеми или практически всеми возрастными группами. Она способна к самоподдержанию вегетативным или семенным путями. Нормальной полночленной называют популяцию, которая состоит из особей всех возрастных групп. Если особи каких-либо возрастных состояний отсутствуют, например проростки или сенильные, то такая ценопопуляция называется нормальной неполночленной. Нормальные неполночленные ценопопуляции имеют монокарпики. Ценопопуляцию, не содержащую молодых особей, а представленную сенильными и субсенильными или даже цветущими особями, но не образующими семян, называют регрессивной. Регрессивная ценопопуляция не способна к самоподдержанию и зависит от заноса зачатков извне.
|
|
|
Возрастная структура ценопопуляции в значительной степени определяется биологическими особенностями вида: периодичностью плодоношения, числом продуцируемых семян и вегетативных зачатков, способностью вегетативных зачатков к омоложению, скоростью перехода особей из одного возрастного состояния в другое, длительностью возрастного состояния, способностью образовывать клоны и др.
У многих животных, так же как и у растений, более длительным является предрепродуктивный период. Так, у поденок (Ephemeridae) он продолжается несколько лет из-за длительного развития личинок. Репродуктивный же их возраст не превышает нескольких дней — время размножения взрослых особей. Пострепродуктивный период здесь практически отсутствует. Популяции быстро восстанавливают свою численность, если особи имеют короткий предрепродуктивный период.
В сокращающихся популяциях преобладают старые особи, которые уже не способны интенсивно размножаться. Данная возрастная структура свидетельствует о неблагоприятных условиях. В быстро растущих популяциях преобладают интенсивно размножающиеся молодые особи. В стабильных популяциях это соотношение, как правило, составляет 1:1. При благоприятных условиях в популяции имеются все возрастные группы и поддерживается сравнительно стабильный уровень численности. На возрастной состав популяции помимо общей продолжительности жизни влияют длительность периода размножения, число генераций в сезон, плодовитость и смертность разных возрастных групп. Так, у полевок (Clethrionomys) взрослые особи могут давать потомство 3 раза в год и более, а молодые особи способны размножаться через два—три месяца (рис. 9.5).

Рис. 9.5. Возрастная структура популяции полевки-экономки
на Крайнем Севере (по С. С. Шварцу, 1980);
II — перезимовавшие; I—IV — поколение от перезимовавших; 1/1—2/1 — соответственно первый и второй приплоды от первого поколения; 1/11 — первый приплод от второго поколения
|
|
|
Популяция лосей в любое время года состоит из 10—11 возрастных групп, однако размножаться особи начинают только с пятой возрастной группы.
Еще более сложная картина наблюдается в популяциях растений. К примеру, дубы (Quercus) дают семенную продукцию в течение столетий. И как результат, популяции у них формируются из огромного количества возрастных групп.
Таким образом, следствием правила максимальной рождаемости (плодовитости, воспроизводства) популяции служит правило стабильности ее возрастной структуры: любая естественная популяция стремится к стабильной возрастной структуре, четкому количественному распределению особей по возрастам. Это правило сформулировано А. Лоткой в 1925 г.
Правило А. Лотки приложимо лишь к высшим организмам с возрастной структурой популяций и не имеет свойств универсальности, хотя в более широком биосистемном смысле оно универсально. Правило стабильности возрастной структуры популяций для многих организмов следует дополнить правилом стабильности соотношения полов, если дифференциация по полу вообще существует, что бывает не всегда. В совокупности эти два правила составляют правило стабильности половозрастной структуры популяции.
Для описания возрастной структуры в популяции выделяют возрастные группы, состоящие из организмов одного возраста, и оценивают численность каждой из этих групп. Результаты представляют в виде диаграмм или пирамиды (рис. 9.6).
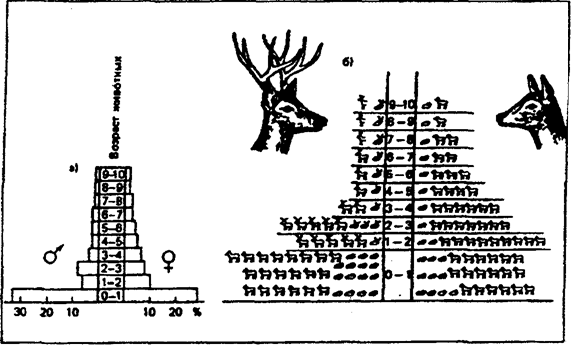
Рис. 9.6. Пирамида возрастов одного из американских оленей:
а) — классическое изображение; б) — изображение с учетом погибших особей в каждой группе (на рисунке слева — самцы, справа — самки) (по Н. Ф. Реймерсу, 1990)
Если в популяции размножение происходит постоянно, то по возрастной структуре устанавливают, сокращается или увеличивается численность. Если основание пирамиды широкое (рис. 9.6), это означает, что рождается больше потомства, рождаемость превышает смертность и численность растет (в данном случае — населения бывшего СССР, 1970 г., и Кении, 1969). Если же особей младших возрастных групп меньше, чем более старых, то численность будет сокращаться (рис.9.7).
|
|
|
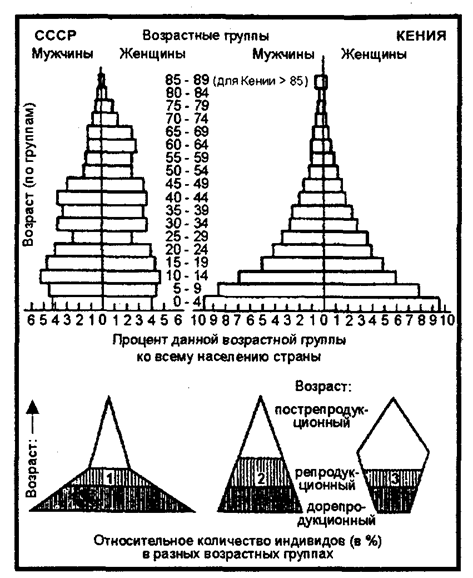
Рис. 9.7. Возрастные пирамиды населения бывшего СССР (1970 г.)
и Кении (1969 г.) и типы возрастных пирамид:
1 — массовое размножение: 2 — стабильная популяция; 3 — сокращающаяся популяция (по Н. Ф. Реймерсу, 1990)
Условия существования особей разного возраста в популяции часто резко различны. Различна и их смертность: например, личинки многих беспозвоночных и рыб подвержены более высокой смертности, чем взрослые особи.
Половой состав популяции
Генетический механизм определения пола обеспечивает расщепление потомства по полу в отношении 1:1, так называемое соотношение полов. Но из этого не следует, что такое же соотношение характерно для популяции в целом. Сцепленные с полом признаки часто определяют значительные различия в физиологии, экологии и поведении самок и самцов. В силу разной жизнеспособности мужского и женского организмов это первичное соотношение нередко отличается от вторичного и особенно от третичного — характерного для взрослых особей. Так, у человека вторичное соотношение полов составляет 100 девочек на 106 мальчиков, к 16—18 годам это соотношение из-за повышенной мужской смертности выравнивается и к 50 годам составляет 85 мужчин на 100 женщин, а к 80 годам — 50 мужчин на 100 женщин (см. рис. 9.7).
Экологическое и поведенческое различия между особями мужского пола могут быть сильно выражены. Так, самцы комаров семейства Culicidae в отличие от кровососущих самок в имагинальный период или не питаются совсем, или ограничиваются слизыванием росы, или потребляют нектар растений. Если даже образ жизни самцов и самок сходен, то они различаются по многим физиологическим признакам: темпам роста, срокам полового созревания, устойчивости к изменениям температуры, голоданию и т. д.
Вторичное и третичное соотношение полов у животных и растений может колебаться и в очень незначительных пределах у разных видов. Существуют популяции, например у некоторых мух, состоящие исключительно из самок. Из самок состоят популяции ряда партеногенетических видов насекомых и ряда других животных. При этом доля партеногенетических самок в разных популяциях может значительно варьировать. У некоторых видов пол изначально определяется не генетическими, а экологическими факторами. У корнеплода Arisaemajaponica решающим фактором во вторичном определении пола является масса клубней: самые крупные и хорошо развитые клубни дают растения с женскими цветами, а мелкие и слабые — с мужскими цветами.
Наглядно прослеживается влияние условий среды на половую структуру популяций у видов с чередованием половых и партеногенетических поколений. При оптимальной температуре дафнии (Daphnia magna) размножаются партеногенетически, а при повышенной или пониженной температуре в популяциях появляются самцы. Появление обоеполого поколения у тли может происходить из-за длины светового дня, температуры, увеличения плотности населения и других факторов.
|
|
|
|
|
|
