 |


Генетический код и его свойства
|
|
|
|
Генетический код — система записи информации о последовательности аминокислот в полипептиде последовательностью нуклеотидов ДНК или РНК. В настоящее время эта система записи считается расшифрованной.
Свойства генетического кода:
- триплетность: каждая аминокислота кодируется сочетанием из трех нуклеотидов (триплетом, кодоном);
- однозначность (специфичность): триплет соответствует только одной аминокислоте;
- вырожденность (избыточность): аминокислоты могут кодироваться несколькими (до шести) кодонами;
- универсальность: система кодирования аминокислот одинакова у всех организмов Земли;
- неперекрываемость: последовательность нуклеотидов имеет рамку считывания по 3 нуклеотида, один и тот же нуклеотид не может быть в составе двух триплетов;
- из 64 кодовых триплетов 61 — кодирующие, кодируют аминокислоты, а 3 — бессмысленные (в РНК — УАА, УГА, УАГ), не кодируют аминокислоты. Они называются кодонами-терминаторами, поскольку блокируют синтез полипептида во время трансляции. Кроме того, есть кодон-инициатор (в РНК — АУГ), с которого трансляция начинается.
Реакции матричного синтеза
Это особая категория химических реакций, происходящих в клетках живых организмов. Во время этих реакций происходит синтез полимерных молекул по плану, заложенному в структуре других полимерных молекул-матриц. На одной матрице может быть синтезировано неограниченное количество молекул-копий. К этой категории реакций относятся репликация, транскрипция, трансляция и обратная транскрипция.
| Название реакции матричного синтеза | Характеристика процесса | Основные компоненты |
| Репликация | Синтез ДНК на матрице ДНК | Дезоксирибонуклеозидтрифосфаты, ферменты |
| Транскрипция | Синтез РНК на матрице ДНК | Участок ДНК, рибонуклеозидтрифосфаты, ферменты |
| Трансляция | Синтез полипептида на матрице РНК | Рибосомы, иРНК, аминокислоты, тРНК, АТФ, ГТФ, ферменты |
| Обратная транскрипция | Синтез ДНК на матрице РНК | Дезоксирибонуклеозидтрифосфаты, ферменты |
Строение гена эукариот
|
|
|
Ген — участок молекулы ДНК, кодирующий первичную последовательность аминокислот в полипептиде или последовательность нуклеотидов в молекулах транспортных и рибосомных РНК. ДНК одной хромосомы может содержать несколько тысяч генов, которые располагаются в линейном порядке. Место гена в определенном участке хромосомы называется локусом. Особенностями строения гена эукариот являются: 1) наличие достаточно большого количества регуляторных блоков, 2) мозаичность (чередование кодирующих участков с некодирующими). Экзоны (Э) — участки гена, несущие информацию о строении полипептида. Интроны (И) — участки гена, не несущие информацию о строении полипептида. Число экзонов и интронов различных генов разное; экзоны чередуются с интронами, общая длина последних может превышать длину экзонов в два и более раз. Перед первым экзоном и после последнего экзона находятся нуклеотидные последовательности, называемые соответственно лидерной (ЛП) и трейлерной последовательностью (ТП). Лидерная и трейлерная последовательности, экзоны и интроны образуют единицу транскрипции. Промотор (П) — участок гена, к которому присоединяется фермент РНК-полимераза, представляет собой особое сочетание нуклеотидов. Перед единицей транскрипции, после нее, иногда в интронах находятся регуляторные элементы (РЭ), к которым относятся энхансеры и сайленсеры. Энхансеры ускоряют транскрипцию, сайленсеры тормозят ее.

Транскрипция у эукариот
Транскрипция — синтез РНК на матрице ДНК. Осуществляется ферментом РНК-полимеразой.
|
|
|
РНК-полимераза может присоединиться только к промотору, который находится на 3'-конце матричной цепи ДНК, и двигаться только от 3'- к 5'-концу этой матричной цепи ДНК. Синтез РНК происходит на одной из двух цепочек ДНК в соответствии с принципами комплементарности и антипараллельности. Строительным материалом и источником энергии для транскрипции являются рибонуклеозидтрифосфаты (АТФ, УТФ, ГТФ, ЦТФ).
В результате транскрипции образуется «незрелая» иРНК (про-иРНК), которая проходит стадию созревания или процессинга. Процессинг включает в себя: 1) КЭПирование 5'-конца, 2) полиаденилирование 3'-конца (присоединение нескольких десятков адениловых нуклеотидов), 3) сплайсинг (вырезание интронов и сшивание экзонов). В зрелой иРНК выделяют КЭП, транслируемую область (сшитые в одно целое экзоны), нетранслируемые области (НТО) и полиадениловый «хвост».

Транслируемая область начинается кодоном-инициатором, заканчивается кодонами-терминаторами. НТО содержат информацию, определяющую поведение РНК в клетке: срок «жизни», активность, локализацию.
Транскрипция и процессинг происходят в клеточном ядре. Зрелая иРНК приобретает определенную пространственную конформацию, окружается белками и в таком виде через ядерные поры транспортируется к рибосомам; иРНК эукариот, как правило, моноцистронны (кодируют только одну полипептидную цепь).
Трансляция
Трансляция — синтез полипептидной цепи на матрице иРНК.
Органоиды, обеспечивающие трансляцию, — рибосомы. У эукариот рибосомы находятся в некоторых органоидах — митохондриях и пластидах (70S-рибосомы), в свободном виде в цитоплазме (80S-рибосомы) и на мембранах эндоплазматической сети (80S-рибосомы). Таким образом, синтез белковых молекул может происходить в цитоплазме, на шероховатой эндоплазматической сети, в митохондриях и пластидах. В цитоплазме синтезируются белки для собственных нужд клетки; белки, синтезируемые на ЭПС, транспортируются по ее каналам в комплекс Гольджи и выводятся из клетки. В рибосоме выделяют малую и большую субъединицы. Малая субъединица рибосомы отвечает за генетические, декодирующие функции; большая — за биохимические, ферментативные.
В малой субъединице рибосомы расположен функциональный центр (ФЦР) с двумя участками — пептидильным (Р-участок) и аминоацильным (А-участок). В ФЦР может находиться шесть нуклеотидов иРНК, три — в пептидильном и три — в аминоацильном участках.
|
|
|
Для транспорта аминокислот к рибосомам используются транспортные РНК, тРНК (лекция №4). Длина тРНК от 75 до 95 нуклеотидных остатков. Они имеют третичную структуру, по форме напоминающую лист клевера. В тРНК различают антикодоновую петлю и акцепторный участок. В антикодоновой петле РНК имеется антикодон, комплементарный кодовому триплету определенной аминокислоты, а акцепторный участок на 3'-конце способен с помощью фермента аминоацил-тРНК-синтетазы присоединять именно эту аминокислоту (с затратой АТФ). Таким образом, у каждой аминокислоты есть свои тРНК и свои ферменты, присоединяющие аминокислоту к тРНК.
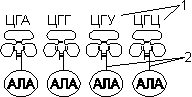
| 
| |
| 1 — антикодон; 2 — участок, связывающий аминокислоту. | Транспорт аминокислот к рибосомам: 1 — фермент; 2 — тРНК; 3 — аминокислота. |
Двадцать видов аминокислот кодируются 61 кодоном, теоретически может быть 61 вид тРНК с соответствующими антикодонами. Но кодируемых аминокислот всего 20 видов, значит, у одной аминокислоты может быть несколько тРНК. Установлено существование нескольких тРНК, способных связываться с одним и тем же кодоном (последний нуклеотид в антикодоне тРНК не всегда важен), поэтому в клетке обнаружено всего около 40 различных тРНК.
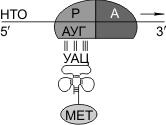
Синтез белка начинается с того момента, когда к 5'-концу иРНК присоединяется малая субъединица рибосомы, в Р-участок которой заходит метиониновая тРНК (транспортирующая аминокислоту метионин). Следует отметить, что любая полипептидная цепь на N-конце сначала имеет метионин, который в дальнейшем чаще всего отщепляется. Синтез полипептида идет от N-конца к С-концу, то есть пептидная связь образуется между карбоксильной группой первой и аминогруппой второй аминокислот.
Затем происходит присоединение большой субъединицы рибосомы, и в А-участок поступает вторая тРНК, чей антикодон комплементарно спаривается с кодоном иРНК, находящимся в А-участке.
|
|
|
Пептидилтрансферазный центр большой субъединицы катализирует образование пептидной связи между метионином и второй аминокислотой. Отдельного фермента, катализирующего образование пептидных связей, не существует. Энергия для образования пептидной связи поставляется за счет гидролиза ГТФ.
Как только образовалась пептидная связь, метиониновая тРНК отсоединяется от метионина, а рибосома передвигается на следующий кодовый триплет иРНК, который оказывается в А-участке рибосомы, а метиониновая тРНК выталкивается в цитоплазму. На один цикл расходуется 2 молекулы ГТФ. В А-участок заходит третья тРНК, и образуется пептидная связь между второй и третьей аминокислотами.
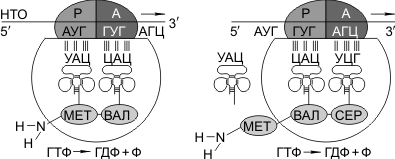
Трансляция идет до тех пор, пока в А-участок не попадает кодон-терминатор (УАА, УАГ или УГА), с которым связывается особый белковый фактор освобождения. Полипептидная цепь отделяется от тРНК и покидает рибосому. Происходит диссоциация, разъединение субъединиц рибосомы.
Скорость передвижения рибосомы по иРНК — 5–6 триплетов в секунду, на синтез белковой молекулы, состоящей из сотен аминокислотных остатков, клетке требуется несколько минут. Первым белком, синтезированным искусственно, был инсулин, состоящий из 51 аминокислотного остатка. Потребовалось провести 5000 операций, в работе в течение трех лет принимали участие 10 человек.
В трансляции можно выделить три стадии: а) инициации (образование иницаторного комплекса), б) элонгации (непосредственно «конвейер», соединение аминокислот друг с другом), в) терминации (образование терминирующего комплекса).
Митотический цикл. Митоз
Митоз — основной способ деления эукариотических клеток, при котором сначала происходит удвоение, а затем равномерное распределение между дочерними клетками наследственного материала.
Митоз представляет собой непрерывный процесс, в котором выделяют четыре фазы: профазу, метафазу, анафазу и телофазу. Перед митозом происходит подготовка клетки к делению, или интерфаза. Период подготовки клетки к митозу и собственно митоз вместе составляют митотический цикл. Ниже приводится краткая характеристика фаз цикла.
Интерфаза состоит из трех периодов: пресинтетического, или постмитотического, — G1, синтетического — S, постсинтетического, или премитотического, — G2.
Пресинтетический период (2 n 2 c, где n — число хромосом, с — число молекул ДНК) — рост клетки, активизация процессов биологического синтеза, подготовка к следующему периоду.
Синтетический период (2 n 4 c) — репликация ДНК.
Постсинтетический период (2 n 4 c) — подготовка клетки к митозу, синтез и накопление белков и энергии для предстоящего деления, увеличение количества органоидов, удвоение центриолей.
|
|
|
Профаза (2 n 4 c) — демонтаж ядерных мембран, расхождение центриолей к разным полюсам клетки, формирование нитей веретена деления, «исчезновение» ядрышек, конденсация двухроматидных хромосом.
Метафаза (2 n 4 c) — выстраивание максимально конденсированных двухроматидных хромосом в экваториальной плоскости клетки (метафазная пластинка), прикрепление нитей веретена деления одним концом к центриолям, другим — к центромерам хромосом.
Анафаза (4 n 4 c) — деление двухроматидных хромосом на хроматиды и расхождение этих сестринских хроматид к противоположным полюсам клетки (при этом хроматиды становятся самостоятельными однохроматидными хромосомами).
Телофаза (2 n 2 c в каждой дочерней клетке) — деконденсация хромосом, образование вокруг каждой группы хромосом ядерных мембран, распад нитей веретена деления, появление ядрышка, деление цитоплазмы (цитотомия). Цитотомия в животных клетках происходит за счет борозды деления, в растительных клетках — за счет клеточной пластинки.
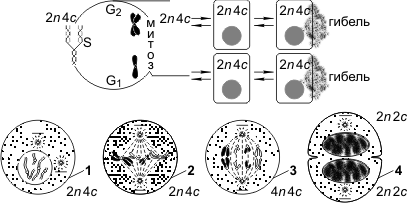
Митотический цикл, митоз: 1 — профаза; 2 — метафаза; 3 — анафаза; 4 — телофаза.
Биологическое значение митоза. Образовавшиеся в результате этого способа деления дочерние клетки являются генетически идентичными материнской. Митоз обеспечивает постоянство хромосомного набора в ряду поколений клеток. Лежит в основе таких процессов, как рост, регенерация, бесполое размножение и др.
Мейоз
Мейоз — это особый способ деления эукариотических клеток, в результате которого происходит переход клеток из диплоидного состояния в гаплоидное. Мейоз состоит из двух последовательных делений, которым предшествует однократная репликация ДНК.
Первое мейотическое деление (мейоз 1) называется редукционным, поскольку именно во время этого деления происходит уменьшение числа хромосом вдвое: из одной диплоидной клетки (2 n 4 c) образуются две гаплоидные (1 n 2 c).
Интерфаза 1 (в начале — 2 n 2 c, в конце — 2 n 4 c) — синтез и накопление веществ и энергии, необходимых для осуществления обоих делений, увеличение размеров клетки и числа органоидов, удвоение центриолей, репликация ДНК, которая завершается в профазе 1.
Профаза 1 (2 n 4 c) — демонтаж ядерных мембран, расхождение центриолей к разным полюсам клетки, формирование нитей веретена деления, «исчезновение» ядрышек, конденсация двухроматидных хромосом, конъюгация гомологичных хромосом и кроссинговер. Конъюгация — процесс сближения и переплетения гомологичных хромосом. Пару конъюгирующих гомологичных хромосом называют бивалентом. Кроссинговер — процесс обмена гомологичными участками между гомологичными хромосомами.
Профаза 1 подразделяется на стадии: лептотена (завершение репликации ДНК), зиготена (конъюгация гомологичных хромосом, образование бивалентов), пахитена (кроссинговер, перекомбинация генов), диплотена (выявление хиазм, 1 блок овогенеза у человека), диакинез (терминализация хиазм).

Мейоз: 1 — лептотена; 2 — зиготена; 3 — пахитена; 4 — диплотена; 5 — диакинез; 6 — метафаза 1; 7 — анафаза 1; 8 — телофаза 1;
9 — профаза 2; 10 — метафаза 2; 11 — анафаза 2; 12 — телофаза 2.
Метафаза 1 (2 n 4 c) — выстраивание бивалентов в экваториальной плоскости клетки, прикрепление нитей веретена деления одним концом к центриолям, другим — к центромерам хромосом.
Анафаза 1 (2 n 4 c) — случайное независимое расхождение двухроматидных хромосом к противоположным полюсам клетки (из каждой пары гомологичных хромосом одна хромосома отходит к одному полюсу, другая — к другому), перекомбинация хромосом.
Телофаза 1 (1 n 2 c в каждой клетке) — образование ядерных мембран вокруг групп двухроматидных хромосом, деление цитоплазмы. У многих растений клетка из анафазы 1 сразу же переходит в профазу 2.
Второе мейотическое деление (мейоз 2) называется эквационным.
Интерфаза 2, или интеркинез (1n 2c), представляет собой короткий перерыв между первым и вторым мейотическими делениями, во время которого не происходит репликация ДНК. Характерна для животных клеток.
Профаза 2 (1 n 2 c) — демонтаж ядерных мембран, расхождение центриолей к разным полюсам клетки, формирование нитей веретена деления.
Метафаза 2 (1 n 2 c) — выстраивание двухроматидных хромосом в экваториальной плоскости клетки (метафазная пластинка), прикрепление нитей веретена деления одним концом к центриолям, другим — к центромерам хромосом; 2 блок овогенеза у человека.
Анафаза 2 (2 n 2 с) — деление двухроматидных хромосом на хроматиды и расхождение этих сестринских хроматид к противоположным полюсам клетки (при этом хроматиды становятся самостоятельными однохроматидными хромосомами), перекомбинация хромосом.
Телофаза 2 (1 n 1 c в каждой клетке) — деконденсация хромосом, образование вокруг каждой группы хромосом ядерных мембран, распад нитей веретена деления, появление ядрышка, деление цитоплазмы (цитотомия) с образованием в итоге четырех гаплоидных клеток.
Биологическое значение мейоза. Мейоз является центральным событием гаметогенеза у животных и спорогенеза у растений. Являясь основой комбинативной изменчивости, мейоз обеспечивает генетическое разнообразие гамет.
Амитоз
Амитоз — прямое деление интерфазного ядра путем перетяжки без образования хромосом, вне митотического цикла. Описан для стареющих, патологически измененных и обреченных на гибель клеток. После амитоза клетка не способна вернуться в нормальный митотический цикл.
Клеточный цикл
Клеточный цикл — жизнь клетки от момента ее появления до деления или смерти. Обязательным компонентом клеточного цикла является митотический цикл, который включает в себя период подготовки к делению и собственно митоз. Кроме этого, в жизненном цикле имеются периоды покоя, во время которых клетка выполняет свойственные ей функции и избирает дальнейшую судьбу: гибель или возврат в митотический цикл.
Основные понятия генетики
Генетика — наука о закономерностях наследственности и изменчивости. Датой «рождения» генетики можно считать 1900 год, когда Г. Де Фриз в Голландии, К. Корренс в Германии и Э. Чермак в Австрии независимо друг от друга «переоткрыли» законы наследования признаков, установленные Г. Менделем еще в 1865 году.
Наследственность — свойство организмов передавать свои признаки от одного поколения к другому.
Изменчивость — свойство организмов приобретать новые по сравнению с родителями признаки. В широком смысле под изменчивостью понимают различия между особями одного вида.
Признак — любая особенность строения, любое свойство организма. Развитие признака зависит как от присутствия других генов, так и от условий среды, формирование признаков происходит в ходе индивидуального развития особей. Поэтому каждая отдельно взятая особь обладает набором признаков, характерных только для нее.
Фенотип — совокупность всех внешних и внутренних признаков организма.
Ген — функционально неделимая единица генетического материала, участок молекулы ДНК, кодирующий первичную структуру полипептида, молекулы транспортной или рибосомной РНК. В широком смысле ген — участок ДНК, определяющий возможность развития отдельного элементарного признака.
Генотип — совокупность генов организма.
Локус — местоположение гена в хромосоме.
Аллельные гены — гены, расположенные в идентичных локусах гомологичных хромосом.
Гомозигота — организм, имеющий аллельные гены одной молекулярной формы.
Гетерозигота — организм, имеющий аллельные гены разной молекулярной формы; в этом случае один из генов является доминантным, другой — рецессивным.
Рецессивный ген — аллель, определяющий развитие признака только в гомозиготном состоянии; такой признак будет называться рецессивным.
Доминантный ген — аллель, определяющий развитие признака не только в гомозиготном, но и в гетерозиготном состоянии; такой признак будет называться доминантным.
Изменчивость — способность живых организмов приобретать новые признаки и свойства. Благодаря изменчивости, организмы могут приспосабливаться к изменяющимся условиям среды обитания.
Различают две основные формы изменчивости: наследственная и ненаследственная.
Наследственная, или генотипическая, изменчивость — изменения признаков организма, обусловленные изменением генотипа. Она, в свою очередь, подразделяется на комбинативную и мутационную. Комбинативная изменчивость возникает вследствие перекомбинации наследственного материала (генов и хромосом) во время гаметогенеза и полового размножения. Мутационная изменчивость возникает в результате изменения структуры наследственного материала.
Ненаследственная, или фенотипическая, или модификационная, изменчивость — изменения признаков организма, не обусловленные изменением генотипа.
Мутации
Мутации — это стойкие внезапно возникшие изменения структуры наследственного материала на различных уровнях его организации, приводящие к изменению тех или иных признаков организма.
Термин «мутация» введен в науку Де Фризом. Им же создана мутационная теория, основные положения которой не утратили своего значения по сей день.
- Мутации возникают внезапно, скачкообразно, без всяких переходов.
- Мутации наследственны, т.е. стойко передаются из поколения в поколение.
- Мутации не образуют непрерывных рядов, не группируются вокруг среднего типа (как при модификационной изменчивости), они являются качественными изменениями.
- Мутации ненаправленны — мутировать может любой локус, вызывая изменения как незначительных, так и жизненно важных признаков в любом направлении.
- Одни и те же мутации могут возникать повторно.
- Мутации индивидуальны, то есть возникают у отдельных особей.
Процесс возникновения мутаций называют мутагенезом, а факторы среды, вызывающие появление мутаций, — мутагенами.
По типу клеток, в которых мутации произошли, различают: генеративные и соматические мутации.
Генеративные мутации возникают в половых клетках, не влияют на признаки данного организма, проявляются только в следующем поколении.
Соматические мутации возникают в соматических клетках, проявляются у данного организма и не передаются потомству при половом размножении. Сохранить соматические мутации можно только путем бесполого размножения (прежде всего вегетативного).
По адаптивному значению выделяют: полезные, вредные (летальные, полулетальные) и нейтральные мутации. Полезные — повышают жизнеспособность, летальные — вызывают гибель, полулетальные — снижают жизнеспособность, нейтральные — не влияют на жизнеспособность особей. Следует отметить, что одна и та же мутация в одних условиях может быть полезной, а в других — вредной.
По характеру проявления мутации могут быть доминантными и рецессивными. Если доминантная мутация является вредной, то она может вызвать гибель ее обладателя на ранних этапах онтогенеза. Рецессивные мутации не проявляются у гетерозигот, поэтому длительное время сохраняются в популяции в «скрытом» состоянии и образуют резерв наследственной изменчивости. При изменении условий среды обитания носители таких мутаций могут получить преимущество в борьбе за существование.
В зависимости от того, выявлен ли мутаген, вызвавший данную мутацию, или нет, различают индуцированные и спонтанные мутации. Обычно спонтанные мутации возникают естественным путем, индуцированные — вызываются искусственно.
В зависимости от уровня наследственного материала, на котором произошла мутация, выделяют: генные, хромосомные и геномные мутации.
Генные мутации
Генные мутации — изменения структуры генов. Поскольку ген представляет собой участок молекулы ДНК, то генная мутация представляет собой изменения в нуклеотидном составе этого участка. Генные мутации могут происходить в результате: 1) замены одного или нескольких нуклеотидов на другие; 2) вставки нуклеотидов; 3) потери нуклеотидов; 4) удвоения нуклеотидов; 5) изменения порядка чередования нуклеотидов. Эти мутации приводят к изменению аминокислотного состава полипептидной цепи и, следовательно, к изменению функциональной активности белковой молекулы. Благодаря генным мутациям возникают множественные аллели одного и того же гена.
Заболевания, причиной которых являются генные мутации, называются генными (фенилкетонурия, серповидноклеточная анемия, гемофилия и т.д.). Наследование генных болезней подчиняется законам Менделя.
Хромосомные мутации
Это изменения структуры хромосом. Перестройки могут осуществляться как в пределах одной хромосомы — внутрихромосомные мутации (делеция, инверсия, дупликация, инсерция), так и между хромосомами — межхромосомные мутации (транслокация).
Делеция — утрата участка хромосомы (2); инверсия — поворот участка хромосомы на 180° (4, 5); дупликация — удвоение одного и того же участка хромосомы (3); инсерция — перестановка участка (6).
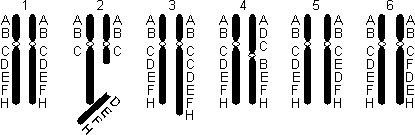
Хромосомные мутации: 1 — парахромосом; 2 — делеция; 3 — дупликация; 4, 5 — инверсия; 6 — инсерция.
Транслокация — перенос участка одной хромосомы или целой хромосомы на другую хромосому.
Заболевания, причиной которых являются хромосомные мутации, относятся к категории хромосомных болезней. К таким заболеваниям относятся синдром «крика кошки» (46, 5р-), транслокационный вариант синдрома Дауна (46, 21 t2121) и др.
Геномные мутации
Геномной мутацией называется изменение числа хромосом. Геномные мутации возникают в результате нарушения нормального хода митоза или мейоза.
Гаплоидия — уменьшение числа полных гаплоидных наборов хромосом.
Полиплоидия — увеличение числа полных гаплоидных наборов хромосом: триплоиды (3 n), тетраплоиды (4 n) и т.д.
Гетероплоидия (анеуплоидия) — некратное увеличение или уменьшение числа хромосом. Чаще всего наблюдается уменьшение или увеличение числа хромосом на одну (реже две и более).
Наиболее вероятной причиной гетероплоидии является нерасхождение какой-либо пары гомологичных хромосом во время мейоза у кого-то из родителей. В этом случае одна из образовавшихся гамет содержит на одну хромосому меньше, а другая — на одну больше. Слияние таких гамет с нормальной гаплоидной гаметой при оплодотворении приводит к образованию зиготы с меньшим или большим числом хромосом по сравнению с диплоидным набором, характерным для данного вида: нулесомия (2 n - 2), моносомия (2 n - 1), трисомия (2 n + 1), тетрасомия (2 n + 2) и т.д.
На генетических схемах, приведенных ниже, показано, что рождение ребенка с синдромом Клайнфельтера или синдромом Тернера-Шерешевского можно объяснить нерасхождением половых хромосом во время анафазы 1 мейоза у матери или у отца.
1) Нерасхождение половых хромосом во время мейоза у матери
| Р | ♀46, XX | × | ♂46, XY | ||
| Типы гамет |  24, XX 24, XX  24, 0 24, 0
| 23, X 23, Y
| |||
| F | 47, XXX трисомия по Х-хромосоме | 47, XXY синдром Клайнфельтера | 45, X0 синдром Тернера- Шерешевского | 45, Y0 гибель зиготы | |
2) Нерасхождение половых хромосом во время мейоза у отца
| Р | ♀46, XX | × | ♂46, XY |
| Типы гамет | 23, X
| 24, XY 22, 0
| |
| F | 47, XXY синдром Клайнфельтера | 45, X0 синдром Тернера- Шерешевского |
Заболевания, причиной которых являются геномные мутации, также относятся к категории хромосомных. Их наследование не подчиняется законам Менделя. Кроме вышеназванных синдромов Клайнфельтера или Тернера-Шерешевского, к таким болезням относятся синдромы Дауна (47, +21), Эдвардса (+18), Патау (47, +15).
Полиплодия характерна для растений. Получение полиплоидов широко используется в селекции растений.
|
|
|
