 |


Филогенез артериальных жаберных дуг
|
|
|
|
В связи с тем что основные артериальные сосуды у млекопитающих и человека формируются на базе закладок жаберных артерий, проследим их эволюцию в филогенетическом ряду позвоночных (рис. 14.30). В эмбриогенезе абсолютного большинства позвоночных закладывается шесть пар артериальных жаберных дуг, соответствующих шести парам висцеральных дуг черепа. В связи с тем что две первые пары висцеральных дуг включаются в состав лицевого черепа, две первые артериальные жаберные дуги быстро редуцируются. Оставшиеся четыре пары функционируют у рыб как жаберные артерии. У наземных позвоночных 3-я пара жаберных артерий теряет связь с корнями спинной аорты и несет кровь к голове, становясь сонными артериями. Сосуды 4-й пары достигают наибольшего развития и вместе с участком корня спинной аорты во взрослом состоянии становятся дугами аорты — основными сосудами большого круга кровообращения.
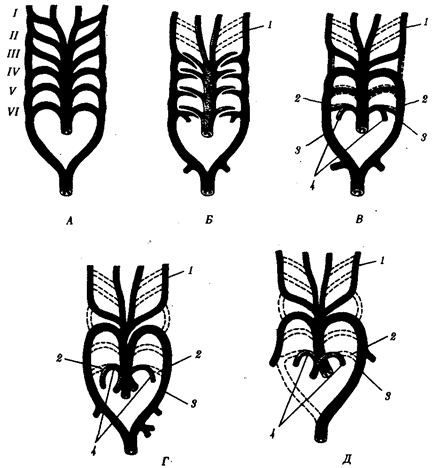
Рис. 14.30. Эволюция артериальных жаберных дуг позвоночных. А— закладка в эмбриогенезе; Б —жаберные дуги рыб; В— жаберные дуги хвостатых земноводных- Г —жаберные дуги пресмыкающихся; Д— жаберные дуги млекопитающих:
I — VI - жаберные дуги; 1 —сонные артерии, 2 —боталлов проток, 3 —артериальные дуги большого круга кровообращения, 4— легочные артерии
У земноводных и пресмыкающихся оба сосуда развиты и принимают участие в кровообращении. У млекопитающих также закладываются оба сосуда 4-й пары, а позже правая дуга аорты редуцируется таким образом, что от нее остается лишь небольшой рудимент— плечеголовной ствол. Пятая пара артериальных дуг в связи с тем, что она функционально дублирует четвертую, редуцируется у всех наземных позвоночных, кроме хвостатых амфибий. Шестая пара, которая снабжает венозной кровью кроме жабр еще и плавательный пузырь, у кистеперых рыб становится легочной артерией.
|
|
|
В эмбриогенезе человека рекапитуляции артериальных жаберных дуг происходят с особенностями: все шесть пар дуг никогда не существуют одновременно. В то время, когда две первые дуги закладываются, а затем перестраиваются, последние пары сосудов еще не начинают формироваться. Кроме того, пятая артериальная дуга уже закладывается в виде рудиментарного сосуда, присоединенного обычно к 4-й паре, и редуцируется очень быстро.
Из атавистических пороков развития сосудов, развивающихся из артериальных жаберных дуг, остановимся на следующих: с частотой 1 случай на 200 вскрытии детей, умерших от врожденных пороков сердца, встречается персистирование обеих дуг аорты 4-й пары. При этом обе дуги, так же как у земноводных или пресмыкающихся, срастаются позади пищевода и трахеи, образуя нисходящую часть спинной аорты. Порок проявляется нарушением глотания и удушьем. Несколько чаще (2,8 случая на 200 вскрытии) встречается нарушение редукции правой дуги аорты с редукцией левой. Эта аномалия часто клинически не проявляется.
Наиболее частый порок (0,5—1,2 случая на 1000 новорожденных) — персистирование артериального, или боталлова, протока (рис. 14.31, А)представляющего собой часть корня спинной аорты между 4-й и'6-и парами артерий слева. Проявляется сбросом артериальной крови из большого круга кровообращения в малый. Очень тяжелый порок развития — персистирование первичного эмбрионального ствола, в результате которого из сердца выходит только один сосуд, располагающийся обычно над дефектом в межжелудочковой перегородке (рис. 14.31, Б). Он обычно заканчивается смертью ребенка. Нарушение дифференцировки первичного эмбрионального ствола может привести к такому пороку развития, как транспозиция сосудов — отхождение аорты от правого желудочка, а легочного ствола — от левого, что встречается в 1 случае на 2500 новорожденных. Этот порок обычно несовместим с жизнью.
|
|
|
Рекапитуляции проявляются и в эмбриональном развитии крупных вен человека. При этом возможно формирование атавистических пороков развития. Среди пороков развития венозного русла укажем на возможность персистирования двух верхних полых вен. Если обе они впадают в правое предсердие, аномалия клинически не проявляется. При впадении левой полой вены в левое предсердие происходит сброс венозной крови в большой круг кровообращения. Иногда обе полые вены впадают в левое предсердие. Такой порок несовместим с жизнью. Данные аномалии встречаются с частотой 1% от всех врожденных пороков сердечно-сосудистой системы.
Очень редкая врожденная аномалия — неразвитие нижней полой вены. Отток крови от нижней части туловища и ног осуществляется в этом случае через коллатерали непарной и полунепарной вен, являющихся рудиментами задних кардинальных вен.
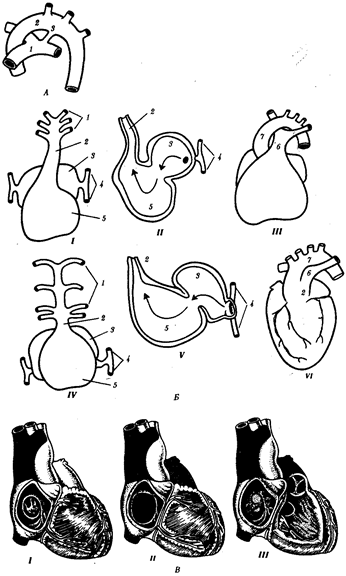
Рис. 14.31. Врожденные пороки развития сердца и сосудов у человека:
А — персистирование артериального (боталлова) протока у человека:
1 —легочный ствол, 2— левая дуга аорты, 3— артериальный (боталлов) проток;
Б— онтофилогенетические основы формирования пороков развития сердца и отходящих от него сосудов у человека: I —сердце зародыша человека в возрасте 5 недель; II —то же, в разрезе III— разделение артериального конуса у зародыша человека в возрасте 6 недель, IV— сердце рыбы, V— то же, в разрезе, VI —персистирование неразделенного артериального конуса у человека: 1— артериальные жаберные дуги, 2— артериальный конус, 3— предсердие, 4— кардинальные вены, 5— желудочек, 6 —общая легочная артерия, 7—левая дуга аорты;
В — аномалии межпредсердной (I, II) и межжелудочковой (III)перегородок
МОЧЕПОЛОВАЯ СИСТЕМА
Выделительная и половая системы в целом выполняют разные функции. Однако их рассматривают в едином комплексе в связи с единством эмбрионального развития и первичной функциональной связью со вторичной полостью тела — целбмом. В эмбриогенезе закладка мочеполовой системы — нефрогонотом — формируется в области ножки сомита в непосредственном контакте с целомом. Половые железы у всех хордовых животных располагаются в целоме. Продукты диссимиляции у всех целоми-ческих животных, в том числе и низших хордовых, также поступают в целом. Наиболее простой путь выведения во внешнюю среду как половых клеток, так и продуктов диссимиляции — через общий канал, начинающийся воронкой в целоме и заканчивающийся выделительной порой на покровах. Таковы взаимоотношения половых желез и выделительных каналов у многих беспозвоночных и, вероятно, у древних предков хордовых (рис. 14.32).
|
|
|
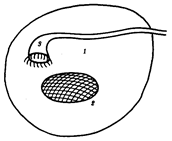
Рис. 14.32. Взаимоотношения целома (1), половой железы (2) и общего выделительного канала (3)
Эволюция почки
Органами выделения позвоночных являются почки — парные компактные органы, структурная единица которых представлена нефроном. В наиболее примитивном виде это — воронка, открывающаяся в целом и соединенная с выделительным канальцем, который впадает в общий выводной проток — мочеточник. В филогенезе позвоночных почка прошла три этапа эволюции: предпочка — головная, или пронефрос; первичная почка—туловищная, или мезонефрос, и вторичная почка - тазовая, или метанефрос.
Предпочка полностью развивается и функционирует как самостоятельный орган у личинок рыб и земноводных. Она находится на переднем конце тела, состоит из 2—12 нефронов, воронки которых открыты в целом, а выводные канальцы впадают в пронефрический канал, который соединен с клоакой. Предпочка имеет сегментарное строение. Продукты диссимиляции фильтруются в целом из кровеносных сосудов, которые поблизости от нефронов формируют клубочки (рис. 14.33, А).
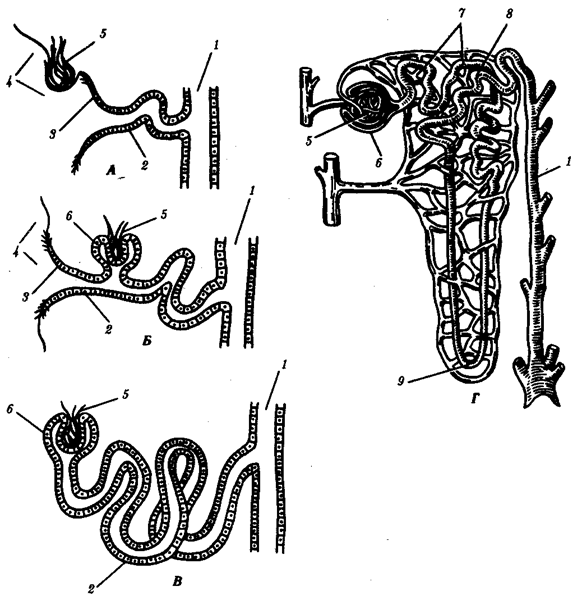
Рис. 14.33. Эволюция тефрона. А— предпочка; Б, В— первичная почка; Г— вторичная почка:
1 —собирательная трубочка, 2— выделительный канадец, 3— нефростом, 4— целом, 5— капиллярный клубочек, 6— капсула, 7, 8— извитой канадец, 9— петля нефрона
У взрослых рыб и земноводных кзади от предпочек, в туловищных сегментах тела, формируются первичные почки, содержащие до нескольких сотен нефронов. В ходе онтогенеза нефроны увеличиваются в количестве за счет их почкования друг от друга с последующей дифференцировкой. Они вступают в связь с кровеносной системой, формируя капсулы почечных клубочков. Капсулы имеют вид двустенных чаш, в которых располагаются сосудистые клубочки, благодаря чему продукты диссимиляции могут поступать из крови непосредственно в нефрон. Некоторые нефроны первичной почки сохраняют связь с целомом через воронки, другие — утрачивают её (рис. 14.33, Б, В).
|
|
|
Выделительные канальцы удлиняются и в них осуществляется обратное всасывание в кровь воды, глюкозы и других веществ, в связи с чем концентрация продуктов диссимиляции в моче повышается. Однако воды с мочой теряется много, поэтому животные, обладающие такой почкой, могут обитать только в водной или влажной среде. Первичная почка сохраняет признаки метамерного строения.
У пресмыкающихся и млекопитающих возникают вторичные почки. Они закладываются в тазовом отделе тела и содержат сотни тысяч нефронов наиболее совершенного строения. У новорожденного ребенка в почке их насчитывается около 1 млн. Они образуются за счет многократного ветвления развивающихся нефронов. Нефроны не имеют воронки и, таким образом, теряют полностью связь с целомом. Канадец нефрона удлиняется, теснее контактирует с кровеносной системой, а у млекопитающих дифференцируется на проксимальный и дистальный участки, между которыми появляется еще и так называемая петля Генле (рис. 14.33, Г).
Такое строение нефрона обеспечивает не только полноценную фильтрацию плазмы крови в капсуле, но и, что более важно, эффективное обратное всасывание в кровь воды, глюкозы, гормонов, солей и других необходимых организму веществ. В результате концентрация продуктов диссимиляции в моче, выделяемой вторичными почками, велика, а само ее количество — мало. У человека, например, за сутки в капсулах нефронов обеих почек фильтруется около 150 л плазмы крови, а мочи выделяется около 2 л. Это позволяет животным, обладающим вторичными почками, быть более независимыми от водной среды и заселять засушливые участки земли. У пресмыкающихся вторичные почки на протяжении всей жизни сохраняются на месте их первоначальной закладки — в тазовой области. В них прослеживаются черты первичного метамерного строения.
Почки млекопитающих располагаются в поясничной области, и у большинства из них внешняя сегментация не выражена. В онтогенезе человека обнаруживается выраженная рекапитуляция в развитии почки: закладывание вначале про-, затем мезо-, а позже метанефроса. Последний развивается в тазовой области, а затем за счет различий в скоростях роста позвоночника, таза и органов брюшной полости перемещается в поясничную область. У пятинедельного зародыша можно обнаружить сосуществование предпочки, первичной, а также зачатков вторичной почки (рис. 14.34).
|
|
|
На начальных этапах развития почка человека сегментирована. Позже ее поверхность сглаживается и метамерность сохраняется лишь во внутреннем строении в виде почечных пирамид. Пороки развития почек у человека, основанные на их филогенезе, многообразны. Сохранение мезонефроса и одностороннее отсутствие вторичной почки описаны пока только у мышей, хотя в принципе такая аномалия возможна и у человека. Относительно часто встречается сегментированная вторичная почка, имеющая один или даже несколько мочеточников; возможно и полное ее удвоение. Часто наблюдается тазовое расположение почки, связанное с нарушением ее перемещения на 2—4-м месяцах зародышевого развития (рис. 14.35).

Рис. 14.34. Пятинедельный зародыш человека с тремя поколениями почек:
1 —предпочка, 2— первичная почка, 3— вторичная почка
Рис. 14.35. Онтофилогенетически обусловленные пороки развития почек:
1 —удвоение почки, 2— удвоенный мочеточник, 3— тазовая эктопия почки, 4— надпочечники
Эволюция половых желез
Половые железы у всех позвоночных развиваются в виде парных складок части нефрогонотома в области ножки сомита. Половые складки вдаются в полость тела и оказываются подвешенными на брыжейке. Первичные половые клетки обособляются у зародышей очень рано — уже на стадии гаструляции. Вначале они обнаруживаются в составе презумптивной эктодермы головного конца эмбриона, затем попадают в энтодерму, откуда активно перемещаются в половые складки. Здесь дифференцирующийся эпителий половой железы, включающий в себя первичные половые клетки, объединяется с соединительнотканной стромой в виде шнуров. Такая гонада индифферентна в половом отношении и может развиваться в дальнейшем как в семенник, так и в яичник в зависимости от генетических и эпигенетических факторов дифференцировки пола.
У хрящевых рыб строение половых желез самок и самцов в значительной степени сходно. У всех остальных позвоночных яичник всегда имеет фолликулярное строение, т.е. содержит пузырьки — фолликулы, в каждом из которых находится одна будущая яйцеклетка. При созревании яйцеклетки стенка фолликула разрывается, и она поступает вначале в брюшную полость, а затем в яйцевод. Семенники содержат семенные трубочки, которые соединены с семявыносящими каналами, по которым зрелые сперматозоиды выходят во внешнюю среду.
Индифферентность развивающейся половой железы позвоночных называют первичным гермафродитизмом. Он эволюционно связан, вероятно, с гермафродитизмом древних предков позвоночных. Доказательством этого может служить наличие у наиболее примитивных современных позвоночных — круглоротых — таких половых желез, передняя часть которых является яичником, а задняя — семенником. У самок некоторых земноводных на протяжении всей жизни сохраняется рудиментарный участок индифферентной половой железы. Зародыш человека до определенного возраста имеет недифференцированные половые железы, которые в зависимости от различных факторов (см. разд. 6.1.2) становятся либо семенниками, либо яичниками. Нарушение их дифференцировки может привести к возникновению ovotestis, в котором сочетаются элементы семенника и яичника. У детей, имеющих ovotestis, обнаруживаются признаки гермафродитизма и в наружных половых органах.
У всех позвоночных с непостоянной температурой тела половые железы находятся в брюшной полости. У большинства млекопитающих мужские гонады перемещаются через паховой канал в мошонку, где температура всегда несколько ниже. Вероятно, пониженная температура более благоприятна для сперматогенеза, обеспечивая более низкий уровень спонтанных мутаций в созревающих сперматозоидах.
У человека семенники, закладываясь в брюшной полости, перемещаются через паховой канал и к 8-му месяцу внутриутробного развития оказываются в мошонке. У 2,2 % мальчиков пубертатного возраста обнаруживаются различные формы крипторхизма — неопущения яичек, которые при этом обычно недоразвиты, а часть семявыносящих канальцев заменена соединительной тканью. Для предотвращения бесплодия таким мальчикам необходимо хирургическое низведение яичек в мошонку в раннем возрасте.
|
|
|
