 |


Немембранные органоиды (органеллы)
|
|
|
|
Рибосомы - органоиды клетки, осуществляющие биосинтез белка. Рибосомы впервые описал Дж. Пелеидол в 1955 году. Термин «рибосомы» предложен Робертсом (1958). Они представляют собой плотные сферические образования (рис. 8) диаметром около 25-30 нм, состоящие из большой и малой субъединиц (рис. 13). Число рибосом в цитоплазме
жи  вых клеток весьма велико как у прокариот, так и у эукариот. В обычной бактериальной клетке содержится, например, до 10000 рибосом, а в эукариотических клетках их число в несколько раз больше. Синтетически активная клетка может содержать несколько миллионов рибосом, на которые приходится до 5% её сухой массы. Рибосомы служат местом белкового синтеза.
вых клеток весьма велико как у прокариот, так и у эукариот. В обычной бактериальной клетке содержится, например, до 10000 рибосом, а в эукариотических клетках их число в несколько раз больше. Синтетически активная клетка может содержать несколько миллионов рибосом, на которые приходится до 5% её сухой массы. Рибосомы служат местом белкового синтеза.
 Различают 2 основных типа рибосом - эукариотные (с константой седиментации 80S) и прокариотные (с константой 70 S). В митохондриях и хлоропластах содержатся мелкие рибосомы (константа седиментации 55S - 70S), осуществляющие автономный синтез белка. В состав рибосомы входят рРНК (3 молекулы у прокариот, 4 молекулы у эукариот) и белки. Молекулы рРНК составляют 50-63% массы рибосомы и образуют её структурный каркас. Каждый из белков рибосомы представлен в ней одной молекулой, т.е. на одну рибосому приходится несколько десятков разных белков (около 55 - для рибосомы прокариот и около 100 - для рибосомы эукариот). Большинство белков специфически связано с определенными участками рРНК. Некоторые белки - т.н. факторы инициации (начала), элонгации (продолжения) и терминации (окончания) - включаются в состав рибосомы только во время биосинтеза белка.
Различают 2 основных типа рибосом - эукариотные (с константой седиментации 80S) и прокариотные (с константой 70 S). В митохондриях и хлоропластах содержатся мелкие рибосомы (константа седиментации 55S - 70S), осуществляющие автономный синтез белка. В состав рибосомы входят рРНК (3 молекулы у прокариот, 4 молекулы у эукариот) и белки. Молекулы рРНК составляют 50-63% массы рибосомы и образуют её структурный каркас. Каждый из белков рибосомы представлен в ней одной молекулой, т.е. на одну рибосому приходится несколько десятков разных белков (около 55 - для рибосомы прокариот и около 100 - для рибосомы эукариот). Большинство белков специфически связано с определенными участками рРНК. Некоторые белки - т.н. факторы инициации (начала), элонгации (продолжения) и терминации (окончания) - включаются в состав рибосомы только во время биосинтеза белка.
Субъединицы рибосомы находятся в состоянии обратимой диссоциации-ассоциации. Сборка рибосом происходит в начале синтеза полипептида, а по завершении синтеза они вновь распадаются на субъединицы. При низкой концентрации ионов магния преобладает диссоциация субъединиц, повышение концентрации ионов магния ведет к их объединению. Одну молекулу иРНК могут одновременно транслировать несколько рибосом (рис. 14), образующих комплекс - полирибосому (полисому). Количество полирибосом в клетке указывает на интенсивность биосинтеза белка.
|
|
|
Аминокислоты, необходимые для синтеза белков, приносятся транспортными РНК (тРНК), а программа синтеза заключена в информационной, или матричной РНК (иРНК), образуемой в ядре клетки и присоединяющейся к рибосоме. В больших субъединицах предполагается наличие куполоподобной структуры, причем находящиеся в этом районе РНК и полипептиды защищены от действия клеточных ферментов (РНК-аз и протеаз). В рибосоме имеются два участка: аминоацильный (А), к которому присоединяется комплекс транспортной РНК с аминокислотой (аминоацил-тРНК), и пептидильный (П), куда продвигается тРНК после образования пептидной связи между принесенной ею аминокислотой и удлиняющейся полипептидной цепью.

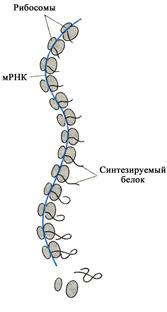 Синтез белка начинается с объединения малой субчастицы рибосомы и инициирующей М-аминоацил-тРНК. Затем к этому комплексу присоединяется большая субчастица, и сформировавшаяся рибосома начинает продвигаться к 3'-концу иРНК. По ходу этого процесса в П-участок рибосомы поступают очередные аминоацил-тРНК в последовательности, определяемой кодонами иРНК. В полипептид включается около
Синтез белка начинается с объединения малой субчастицы рибосомы и инициирующей М-аминоацил-тРНК. Затем к этому комплексу присоединяется большая субчастица, и сформировавшаяся рибосома начинает продвигаться к 3'-концу иРНК. По ходу этого процесса в П-участок рибосомы поступают очередные аминоацил-тРНК в последовательности, определяемой кодонами иРНК. В полипептид включается около
20 аминокислот в 1 секунду; белковая молекула среднего размера синтезируется за
20-60 с. Пока продолжается синтез белка данной рибосомой, новая рибосома занимает
освобождающееся на иРНК место. По этой причине активно транслируемая иРНК находится в полисомах.
Белки, которые после синтеза остаются в гиалоплазме клетки и используются ею далее, обычно синтезируются на свободных полисомах
(рис. 8). Полисомы, которые своими большими субъединицами прикреплены к мембранам ЭПС, синтезируют белки, накапливающиеся в просвете цистерн ЭПС и в дальнейшем либо секретируемые клеткой, либо запасаемые ею внутри гранул (например, лизосомальные ферменты). На полисомах, связанных с мембранами ЭПС, синтезируется также большая часть интегральных мембранных белков.
|
|
|
У эукариот рибосомы образуются в ядрышке. На ядрышковой ДНК синтезируются предшественники рРНК, которые покрываются поступающими из цитоплазмы рибосомальными белками, расщепляются до нужных размеров и формируют рибосомные субчастицы, выходящие в цитоплазму. Поэтому полностью сформированных рибосом в ядре нет. Рибосомная РНК составляет основную массу клеточной РНК. Она обусловливает базофильную окраску ядрышка и участков эргастоплазмы.
Микротрубочки представляют собой полые цилиндрические образования диаметром 24-25 нм и длиной до нескольких микрометров (рис. 8).



|
Стенка микротрубочек толщиной 5 нм состоит из спиралевидно уложенных протофиламентов - нитей диаметром 5 нм (рис. 15). Каждая нить образована димерами из глобулярных белковых молекул α- и β-тубулина.
Микротрубочки формируют лабильную систему, в которой поддерживается динамическое равновесие между их сборкой и распадом (диссоциацией). Большинство микротрубочек имеет закреплённый («-») и свободный («+») концы. Последний обеспечивает удлинение и укорочение трубочек. В образовании микротрубочек путём самосборки участвуют мелкие сферические тельца - сателлиты (центры организации микротрубочек), содержащиеся в клеточном центре и в базальных тельцах ресничек, а также центромеры хромосом. Если полностью разрушить микротрубочки цитоплазмы, то они отрастают от клеточного центра со скоростью 1 мкм/мин. Разрушение микротрубочек приводит к изменению формы клетки (животная клетка обретает обычно сферическую форму). При этом нарушаются структура клетки и распределение органоидов.
В клетке микротрубочки могут располагаться: а) в виде отдельных элементов; б) в пучках, в которых они связаны друг с другом поперечными мостиками (отростки нейронов); в) в составе пар или дублетов (осевая нить ресничек и жгутиков); г) в составе триплетов (центриоли и базальные тельца). В двух последних вариантах микротрубочки частично сливаются друг с другом.
|
|
|
Основными функциями микротрубочек являются: 1) поддержание формы и полярности клетки; 2) обеспечение упорядоченности расположения компонентов клетки; 3) участие в образовании других, более сложных органоидов (центриоли, реснички и т.д.); 4) участие во внутриклеточном транспорте; 5) обеспечение движения хромосом при митотическом делении клетки; 6) обеспечение движения ресничек.

 Микрофиламенты. Микрофиламентами названы тонкие белковые нити диаметром 5-7 нм, встречающиеся практически во всех типах клеток. Они могут располагаться в цитоплазме пучками, сетевидными слоями или поодиночке (рис. 8, 16). Основным белком микрофиламентов является актин, на долю которого приходится до 5% от общего количества белков. Кроме него в состав микрофиламентов могут входить миозин, тропомиозин, а также несколько десятков актинсвязывающих белков. Молекула актина имеет обычно вид двух спирально скрученных нитей. Непосредственно под плазмолеммой располагается кортикальная сеть, в которой микрофиламенты переплетены между собой и соединены друг с другом с помощью особых белков, например, филамина. Кортикальная сеть обусловливает плавность изменения формы клеток, постепенно перестраиваясь с участием актин-расщепляющих ферментов. Тем самым она препятствует резкой и внезапной деформации клетки при механических воздействиях. Отдельные микрофиламенты кортикальной сети прикрепляются к интегральным и трансмембранным белкам плазмолеммы, а также к так называемым адгезионным соединениям (фокальным контактам), которые связывают клетку с компонентами межклеточного вещества или с другими клетками. Микрофиламенты более устойчивы к физическим и химическим воздействиям, чем микротрубочки.
Микрофиламенты. Микрофиламентами названы тонкие белковые нити диаметром 5-7 нм, встречающиеся практически во всех типах клеток. Они могут располагаться в цитоплазме пучками, сетевидными слоями или поодиночке (рис. 8, 16). Основным белком микрофиламентов является актин, на долю которого приходится до 5% от общего количества белков. Кроме него в состав микрофиламентов могут входить миозин, тропомиозин, а также несколько десятков актинсвязывающих белков. Молекула актина имеет обычно вид двух спирально скрученных нитей. Непосредственно под плазмолеммой располагается кортикальная сеть, в которой микрофиламенты переплетены между собой и соединены друг с другом с помощью особых белков, например, филамина. Кортикальная сеть обусловливает плавность изменения формы клеток, постепенно перестраиваясь с участием актин-расщепляющих ферментов. Тем самым она препятствует резкой и внезапной деформации клетки при механических воздействиях. Отдельные микрофиламенты кортикальной сети прикрепляются к интегральным и трансмембранным белкам плазмолеммы, а также к так называемым адгезионным соединениям (фокальным контактам), которые связывают клетку с компонентами межклеточного вещества или с другими клетками. Микрофиламенты более устойчивы к физическим и химическим воздействиям, чем микротрубочки.
Основными функциями микрофиламентов являются: 1) обеспечение определённой жёсткости и упругости клетки за счёт кортикальной сети микрофиламентов; 2) изменение консистенции цитозоля, в том числе при переходе золя в гель; 3) участие в эндоцитозе и экзоцитозе; 4) обеспечение подвижности немышечных клеток (например, нейтрофилов и макрофагов), в основе которой лежит изменение формы клеточной поверхности вследствие регулируемой полимеризации актина; 5) участие в сокращении мышечных клеток и волокон; 6) стабилизация локальных выпячиваний плазматической мембраны, обеспечиваемой пучками поперечно сшитых актиновых филаментов (микроворсинки, стереоцилии); 7) участие в формировании межклеточных соединений (опоясывающие десмосомы и др.).
|
|
|
Органоиды специального значения. Присутствуют только в специализированных клетках отдельных типов. К ним отнесены реснички, жгутики, микроворсинки, микрофибриллы и др.
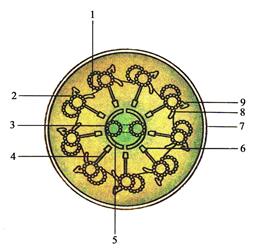
Реснички и жгутики представляют собой выросты цитоплазмы, в которых находится осевая нить, или аксонема (рис. 17, 18). Последняя представляет собой каркас из микротрубочек. Длина ресничек может составлять 2-10 нм, а их количество на поверхности одной клетки достигает нескольких сотен. Длина жгутика изменяется в широких пределах (спермии человека несут один жгутик длиной 50-70 мкм). Аксонема образована 9 периферийными парами микротрубочек и одной парой, расположенной в центре образующегося цилиндра. В каждой периферийной паре из-за частичного слияния микротрубочек одна микротрубочка полная, а вторая неполная, т.к. имеет 2-3 общих димера с первой микротрубочкой (рис. 17). Центральная пара микротрубочек окружена центральной оболочкой. От неё к периферическим парам микротрубочек расходятся в виде лучей так называемые радиальные спицы. Периферические пары связаны между со-
 |
|
|
бой мостиками из белка нексина. Кроме этого, от первой микротрубочки (микротрубочки А) одной пары ко второй микротрубочке (В) соседней пары отходят своеобразные «ручки» из белка динеина. Последний обладает активностью АТФазы.
Колебательные движения жгутиков и биение ресничек обусловлены скольжением соседних дублетов в аксонеме, которое опосредуется движением динеиновых ручек.
|
|
|
В основании каждой реснички или жгутика лежит базальное тельце. Строением оно напоминает центриоль. На уровне апикального конца тельца третья микротрубочка (микротрубочка С) каждого триплета заканчивается, а первая (А) и вторая (В) микротрубочки продолжаются в соответствующие микротрубочки аксонемы жгутика или реснички. В процессе развития жгутика (реснички) базальное тельце служит матрицей, обеспечивающей сборку компонентов аксонемы.

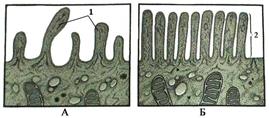 Микроворсинки - это выросты цитоплазмы клетки диаметром 0,1 мкм и длиной 1 мкм (рис. 19). Они многократно увеличивают поверхность клетки, на которой может происходить (например, в тонком кишечнике) расщепление и всасывание веществ. На апикальной поверхности эпителиальной клетки тощей кишки может находиться
Микроворсинки - это выросты цитоплазмы клетки диаметром 0,1 мкм и длиной 1 мкм (рис. 19). Они многократно увеличивают поверхность клетки, на которой может происходить (например, в тонком кишечнике) расщепление и всасывание веществ. На апикальной поверхности эпителиальной клетки тощей кишки может находиться
до нескольких тысяч микроворсинок, которые формируют так
называемую щёточную каёмку
(рис. 19), увеличивающую поверхность клетки более чем в 30 раз.
Каждая микроворсинка имеет внутренний каркас, образованный пучком из примерно 40 микрофиламентов. Пучок ориентирован вдоль продольной оси ворсинки и закреплён в апикальной части микроворсинки особыми белковыми мостиками (молекулами минимиозина), фиксирующимися на внутренней поверхности плазмолеммы. Микрофиламенты пучка соединены между собой поперечными сшивками из белков виллина и фимбрина. В области основания микроворсинки микрофиламенты пучка вплетаются в терминальную сеть, содержащую миозиновые филаменты. Предполагается, что конфигурация и тонус микроворсинок обусловливаются взаимодействием актиновых и миозиновых филаментов.
Промежуточные филаменты представляют собой сплетённые белковыми нитями канаты толщиной около 10 нм. Такой показатель обусловил отведение им промежуточного места между микротрубочками и микрофиламентами. Промежуточные филаменты образуют трёхмерные сети в клетках различных тканей животного организма (рис. 20). Они окружают ядро и могут находиться в различных участках цитоплазмы, образуют межклеточные соединения (десмосомы и полудесмосомы), располагаются внутри отростков нервных клеток. Основными функциями промежуточных филаментов являются структурная и опорная, а также функция распределения органелл в определённых участках клетки.

 Включения клетки. Представляют собой непостоянные компоненты цитоплазмы, образованные отложениями веществ, временно выведенными из обмена, или его конечными продуктами. Выделяют трофические, секреторные, экскреторные, специфические и пигментные включения. К трофическим клеточным включениям относятся те, которые отражают метаболизм клетки (липиды, белок, гликоген). В связи с этим трофические включения подразделяют на липидные (жировые), белковые и углеводные
Включения клетки. Представляют собой непостоянные компоненты цитоплазмы, образованные отложениями веществ, временно выведенными из обмена, или его конечными продуктами. Выделяют трофические, секреторные, экскреторные, специфические и пигментные включения. К трофическим клеточным включениям относятся те, которые отражают метаболизм клетки (липиды, белок, гликоген). В связи с этим трофические включения подразделяют на липидные (жировые), белковые и углеводные
(рис. 8). Липидные включения встречаются в виде мелких или крупных липидных капель. Последние образуются в результате слияния первых. Липидные включения могут служить энергетическим субстратом. Из углеводных включений чаще всего встречаются зёрна крахмала и гранулы гликогена (рис. 8). Примером белковых трофических включений являются включения яйцеклеток, входящие в состав желтка. Секреторные клеточные включения характерны для железистых клеток. Секреторные включения представлены обычно мембранными пузырьками, заполненными секретируемым клеткой веществом. Экскреторные включения сходны с секреторными, однако в отличие от последних, содержат вредные продукты метаболизма, подлежащие удалению из клетки. Специфическими включениями обладают высокоспециализированные клетки и клетки со специфическим метаболизмом. Специфическим клеточным включением эритроцитов является гемоглобин - дыхательный пигмент. К пигментным включениям относятся меланин, гемоцианин и др. Наиболее распространены трофические клеточные включения - капли жира, глыбки гликогена, желток яйцеклеток.
Ядерный аппарат клетки
Ядро - главный обязательный органоид клетки у многоклеточных и многих одноклеточных организмов. Впервые ядро в яйцеклетке курицы наблюдал Я. Пуркине в 1825 году. В растительных клетках ядро описано
Р. Броуном (1831-1833), в животных клетках - Т. Шванном (1838-1839).
По характеру организации ядерного аппарата живые организмы разделяются на две группы (надцарства): прокариотические и эукариотические. У эукариот ядерный аппарат представлен клеточным ядром, а у прокариот - кольцевой молекулой ДНК.
Биологическое значение ядра определяется его главным содержимым - гигантскими молекулами ДНК, способными к транскрипции и трансляции. Основными функциями ядра клетки являются: 1) хранение генетической (наследственной) информации клетки; 2) обеспечение реализации генетической информации, контролирующей разнообразные процессы в клетке; 3) воспроизведение и передача генетической информации при делении клетки.
Клетка содержит, как правило, одно ядро, однако существуют и многоядерные клетки. Последние образуются при незавершающемся делении клеток (отсутствует цитотомия или разделение цитоплазмы) либо в результате слияния нескольких одноядерных клеток.
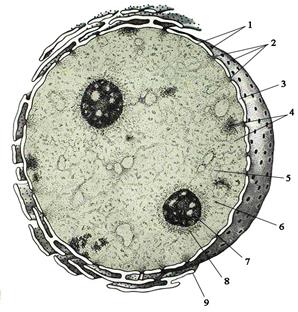
Ядро обычно соответствует форме клетки и имеет сферическую форму в сферических или кубических клетках (рис. 8), вытянутую (эллипсовидную) форму в призматических клетках, уплощённую форму в плоских клетках. Однако часто встречаются клетки с бобовидным, палочковидным, многолопастным и сегментированным ядрами. Варьирует и расположение ядра в клетке: оно может располагаться в центре клетки, вблизи её базального (основного) полюса и на периферии под плазмолеммой. Несмотря на постоянство размера ядра для определённого типа клеток, оно может увеличиваться при усилении активности функций клетки и, наоборот, уменьшаться при её ослаблении.
Ядро интерфазной (неделящейся) клетки состоит из 4-х компонентов (рис. 21): кариоплазмы (ядерного сока или кариолимфы), хроматина, ядрышка и кариолеммы (ядерной оболочки, или нуклеолеммы). Кариоплазма представляет собой жидкий компонент ядра, состоящий из воды и растворённых (или взвешенных) в ней веществ (РНК, ферменты, гликопротеины, ионы и т.п.). Аналогично гиалоплазме она является бесструктурной фазой, которая создаёт для хроматина и ядрышка специфическое микроокружение, обеспечивающее их нормальное функционирование.
Хроматин имеет вид мелких зёрнышек и глыбок, окрашиваемых основными красителями. Он состоит из ДНК и белков. Хроматин являет-
 |
|
|
ся, по сути, интерфазным состоянием хромосом и представляет собой длинные и тонкие перекрученные нити, степень спирализации которых различается по их длине. Последнее обстоятельство послужило основанием для выделения двух разновидностей хроматина: эухроматина и гетерохроматина (рис. 21).
Эухроматин представляет собой деспирализованные (раскрученные) и участвующие в транскрипции сегменты хромосом. Он плохо окрашивается и практически не виден в световой микроскоп.
Гетерохроматин - это спирализованные (плотно скрученные) сегменты хромосом, лишённые активности (не обладающие способностью к транскрипции). Гетерохроматин хорошо окрашивается основными красителями и при наблюдении с помощью светового микроскопа имеет вид гранул и глыбок. Основные скопления гетерохроматина располагаются по периферии ядра, а также вокруг ядрышек. Более мелкие глыбки хроматина разбросаны по всему ядру. Скопление гетерохроматина, соответствующее одной (полностью спирализованной и неактивной) Х-хромосоме у особей женского пола, получило название тельца Барра. Последнее располагается обычно под кариолеммой, а в зернистых лейкоцитах крови имеет вид маленькой добавочной дольки ядра - «барабанной палочки».
При повышении синтетической активности клетки, предполагающей интенсификацию процессов транскрипции, количество эухроматина увеличивается, а содержание гетерохроматина уменьшается. Снижение синтетической активности клетки выражается в увеличении содержания гетерохроматина. Полное угнетение функции ядра (например, в повреждённых и гибнущих клетках, в клетках наружных слоёв эпидермиса) ведёт к уменьшению его размеров, исчезновению эухроматина, интенсивному и равномерному окрашиванию основными красителями (явление кариопикноза).
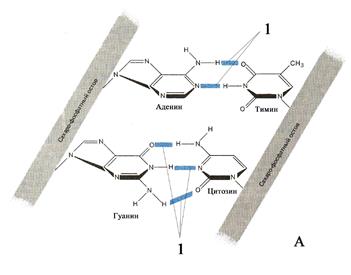
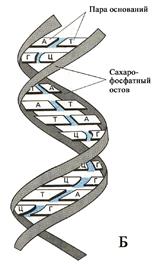
Поскольку полностью деспирализованная молекула ДНК (рис. 22) имеет в среднем длину около 5 см, то расположение в ядре диаметром до
10 мкм предполагает её специфическую упаковку. Роль последней становится более очевидной с учётом того, что общая длина молекул ДНК всех клеток одного человека превышает диаметр Солнечной системы. В клеточном ядре компактная упаковка ДНК осуществляется посредством гистоновых (основных) белков. Она обеспечивает как упорядоченность расположения длинных молекул ДНК в маленьком объёме ядра, так и контроль функциональной активности генов. Упаковка ДНК включает несколько уровней.
Первый уровень заключается в образовании нуклеосомной нити: двойная спираль ДНК наматывается на октамер дисковидной формы, как бы дважды обвивая его, и уходит к очередному октамеру. Октамер состоит из 8 глобулярных молекул гистонов (по 2 молекулы каждого из 4-х ти-

|

|
пов белков: Н2А, Н2В, Н3, Н4). Обвитый двумя витками молекулы ДНК октамер назван нуклеосомой (рис. 23). Нуклеосомы разделены короткими участками связывающей (лигандной) ДНК. ДНК хроматина связана также с негистоновыми белками, которые регулируют избирательную активность генов. В такой регуляции могут участвовать и гистоны, ограничивая доступность ДНК для других ДНК-связывающих белков.
Второй уровень упаковки приводит к скручиванию нуклеосомной нити в спираль диаметром 30-36 нм, именуемую хроматиновой фибриллой. В период между клеточными делениями каждая хромосома представлена двумя хроматиновыми фибриллами (рис. 24).
Третий уровень упаковки ДНК сводится к образованию хроматиновыми фибриллами петель (петельных доменов) диаметром около 300 нм (рис. 24).
Четвертый уровень заключается в более плотной упаковке петельных доменов в так называемые «конденсированные участки хромосом» диаметром около 700 нм. Последние являются непосредственной частью метафазной хромосомы (хромосомы, находящейся в метафазе митотического деления) толщиной примерно 1400 нм (рис. 24).
 Ядрышко выявляется при световой микроскопии как мелкая плотная гранула диаметром 1-3 мкм. Ядрышко отличается высокой концентрацией рибонуклеопротеида, в связи с чем интенсивно окрашивается основными красителями. С повышением функциональной активности клетки увеличиваются как размеры, так и количество ядрышек. Наиболее крупные ядрышки характерны для эмбриональных и активно синтезирующих белки клеток, а также для клеток быстрорастущих злокачественных опухолей.
Ядрышко выявляется при световой микроскопии как мелкая плотная гранула диаметром 1-3 мкм. Ядрышко отличается высокой концентрацией рибонуклеопротеида, в связи с чем интенсивно окрашивается основными красителями. С повышением функциональной активности клетки увеличиваются как размеры, так и количество ядрышек. Наиболее крупные ядрышки характерны для эмбриональных и активно синтезирующих белки клеток, а также для клеток быстрорастущих злокачественных опухолей.
 Ядрышко формируется участками различных молекул ДНК, кодирующих рибосомную РНК (рРНК). Такие участки ДНК названы «ядрышковыми организаторами» (рис. 25). У человека они находятся в ДНК 13й, 14й, 15й, 21й и 22й пар хромосом. Ядрышко не имеет собственной оболочки (мембраны) и исчезает (диспергируется) в профазе митоза из-за того, что ядрышковые организаторы «растаскиваются» в соответствующие пары формирующихся хромосом. В телофазе митотического деления ядрышки формируются вновь участками деконденсирующихся хромосом.
Ядрышко формируется участками различных молекул ДНК, кодирующих рибосомную РНК (рРНК). Такие участки ДНК названы «ядрышковыми организаторами» (рис. 25). У человека они находятся в ДНК 13й, 14й, 15й, 21й и 22й пар хромосом. Ядрышко не имеет собственной оболочки (мембраны) и исчезает (диспергируется) в профазе митоза из-за того, что ядрышковые организаторы «растаскиваются» в соответствующие пары формирующихся хромосом. В телофазе митотического деления ядрышки формируются вновь участками деконденсирующихся хромосом.
Основной функцией ядрышка является синтез рРНК и сборка её в предшественники субъединиц рибосомы. В процессе транскрипции генов ядрышковых организаторов синтезируется единая крупная молекула рРНК. Последняя связывается с поступившими в ядро из цитоплазмы белками, формируя рибонуклеопротеид (РНП), который затем расщепляется на 3 вида РНК: два из них соединяются с молекулами добавочных белков, образуя предшественники большой субъединицы рибосомы, а третий формирует предшественник малой субъединицы рибосомы. Все предшественники субъединиц поступают через ядерные поры в цитоплазму, где окончательно созревают. Ядрышко обычно окружено перинуклеолярным хроматином, который может частично проникать внутрь его, давая начало интрануклеолярному хроматину.
Кариолемма (ядерная оболочка, нуклеолемма) образуется наружной и внутренней мембранами, разделёнными перинуклеарным пространством шириной 15-40 нм (рис. 21). Обе мембраны смыкаются в области ядерных пор. Наружная мембрана кариолеммы является непосредственным продолжением мембран эндоплазматической сети, при этом перинуклеарное пространство соответствует полости цистерн гранулярной эндоплазматической сети (рис. 21). На поверхности наружной мембраны кариолеммы располагаются рибосомы. Внутренняя мембрана кариолеммы гладкая, её интегральные белки связаны с ядерной пластинкой (ламиной) толщиной 80-300 нм. Последняя образована переплетенными промежуточными филаментами, формирующими своеобразный кариоскелет. Ламина выполняет важные функции в поддержании формы ядра, упорядоченной укладке хроматина, формировании кариолеммы при завершении клеточного деления, а также в структурной организации пор.
От 3 до 35% поверхности кариолеммы приходится на ядерные поры, количество которых может достигать в клетках животных и человека 2000-4000 (рис. 21). Последние отсутствуют только в ядрах сперматозоидов. С возрастанием активности клетки количество ядерных пор увеличивается. Совокупность структур, связанных с ядерной порой, названа комплексом ядерной поры. В области поры округлые края наружной и внутренней ядерных мембран сомкнуты, а снаружи и изнутри кариолеммы параллельно друг другу расположены два кольца диаметром по 80 нм, каждое из которых содержит 8 белковых гранул (рис. 26). От гранул в центр поры
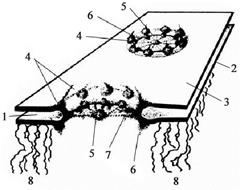 простираются фибриллы толщиной около 5 нм, сходящиеся у центральной гранулы (по мнению некоторых учёных, она представляет собой транс-
простираются фибриллы толщиной около 5 нм, сходящиеся у центральной гранулы (по мнению некоторых учёных, она представляет собой транс-
портируемую субъединицу рибосомы). Через водный канал комплекса ядерной поры диаметром 9 нм проходят ионы и мелкие водорастворимые молекулы. Более крупные молекулы и частицы перемещаются в области  центральной гранулы. Ядерные по-
центральной гранулы. Ядерные по-
ры обеспечивают избирательный транспорт веществ из ядра в цитоплазму и обратно. Особое место в нём занимает перемещение в цитоплазму крупных субъединиц рибосом, предполагающее изменение конформации как субъединицы, так и порового комплекса.
Жизненный цикл клетки
|
|
|
