 |


Поверхностные клетки и клетки крипт специализированы для абсорбции и секреции
|
|
|
|
Абсорбция и секреция солей, приводящие к перемещению воды через стенку тонкой кишки, осуществляются специализированными клетками эпителия. Абсорбция осуществляется эпителиальными клетками ворсинок, которые активно абсорбируют Na+. Секреция производится клетками, расположенными в криптах, которые активно перемещают Cl–. Несмотря на то, что эти клетки активно транспортируют столь разные ионы, параллельно осуществляется и котранспорт: абсорбция Na+ не может происходить без абсорбции Cl–, а секреция Cl– происходит с параллельным перемещением Na+. Активный транспорт является основой перемещения ионов, а перемещение ионов с противоположным зарядом — также неотъемлемая часть этого процесса.
На Рис. 5-6 показаны молекулярные механизмы транспорта солей клетками ворсинок и крипт в тонкой кишке. В обоих типах клеток вектор транспорта ионов связан с выраженным полярным распределением мембранных белков, которые способствуют транспорту ионов через апикальные и базолатеральные участки мембраны клеток. В настоящее время найдены и специальные гены, ответственные за синтез этих белков.
Особой структурой как в абсорбирующих, так и в секретирующих клетках является Na+,K+-ATФaзa. Этот транспортный белок расположен исключительно на базолатеральных участках мембраны клеток кишечника и связывает непосредственно процессы ионного транспорта и энергетические процессы в клетках. Na+,K+-АТФаза поддерживает низкий уровень внутриклеточного Na+ и высокую концентрацию внутриклеточного К+ за счет гидролиза АТФ. Na+,K+-ATФaзa создает градиенты концентраций Na+ и K+, необходимые для поддержания абсорбционной и секреторной активности в эпителиальных клетках кишечника.
|
|
|
Абсорбирующие клетки
Мембрана щеточной каемки абсорбирующих клеток специализирована в отношении пассивного транспорта Na+ в клетки из просвета кишки. В тонкой кишке это происходит преимущественно двумя путями. Первый путь: антипорт Na+/H+, специализирующийся на катализе обмена этих ионов в соотношении 1:1. При этом ионы натрия входят в клетку, а протоны водорода выходят из клетки. Второй механизм — котранспортер Na+/глюкоза, переносящий внутрь клетки ионы Na+ и молекулы глюкозы в соотношении 1:1, либо в соотношении 2:1. Таким образом, в присутствии глюкозы Na+ поступает в клетку вместе с ней. В этом механизме свободная энергия натриевого градиента способствует абсорбции глюкозы. Однако необходимо отметить, что наличие глюкозы в просвете кишки резко повышает абсорб
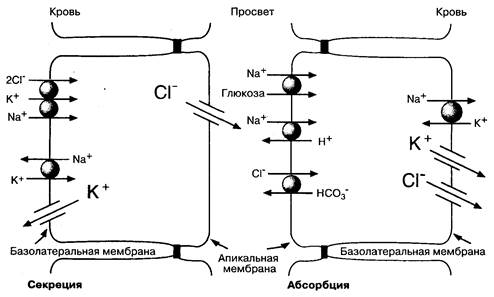
Рис. 5-6. Молекулярные механизмы транспорта веществ клетками ворсинок и крипт в тонкой кишке. Показаны некоторые мембранные белки, участвующие в транспорте солей через апикальные и базолатеральные участки мембраны. Разница электрических потенциалов (Vm) между апикальными и базолатеральными участками мембран абсорбирующих и секретирующих клеток ориентирована таким образом, что клетки относительно экстрацеллюлярной жидкости имеют отрицательный заряд. Величина Vm на базолатеральном участке мембраны больше, чем Vm апикального участка мембраны, поэтому трансэпителиальный электропотснциал, измеренный через слой эпителиальных клеток, является отрицательным со стороны просвета кишки
цию натрия, усиливая его проникновение в клетки. Этот механизм лежит в основе использования растворов глюкозы перорально для регидратации больных с диареей, так как глюкоза повышает абсорбцию Na+ и воды. Растворы, содержащие только необходимые электролиты, менее эффективны, поскольку они не стимулируют глюкозозависимый транспорт Na+ в клетки.
|
|
|
Na+,K+-ATФaзa базолатерального участка мембраны и белки-переносчики Na+ в апикальных отделах мембраны клеток отвечают за трансцеллюлярный перенос Na+ из просвета кишки в кровь; поэтому активный транспорт Na+ — основной механизм абсорбции солей и воды. Абсорбция Сl–, соответствующая абсорбции натрия, происходит двумя путями. Часть Сl– абсорбируется из межклеточного пространства благодаря слабому электрическому потенциалу, отрицательному со стороны просвета кишки. Кроме этого, существует и трансцеллюлярный механизм абсорбции Сl–, обеспечиваемый Сl–/НСО3–-антипортом в апикальном участке мембраны. Механизм транспорта хлора через базолатеральный участок мембраны пока не совсем понятен: это может быть либо Сl–-селективный канал, либо Сl–,K+-котранспортер. Образование Н+ и НСО3– из угольной кислоты с помощью карбоангидразы способствует сочетанному транспорту Na+ и Сl–.
Секреторные клетки
Транспортные белки в секреторных клетках отличаются от транспортных белков в клетках ворсинок (рис. 5-6). Содержание Na+ в клетках регулируется белками базолатерального участка мембраны, в то время как проницаемость для Na+ в апикальных отделах клеток относительно мала. Натрий поступает в клетку через базолатеральный участок мембраны посредством механизма сочетанного транспорта; в каждом цикле в клетку переносится по одному иону Na+ и К+ и по два иона Cl–. Так как суммарный заряд при таком переносе ионов равен нулю, скорость транспорта зависит не от мембранного потенциала, а от концентрации этих трех ионов. Суммарный эффект направленных внутрь градиентов Na+ и Cl– и направленного наружу градиента 1C приводит к перемещению всех трех ионов внутрь клетки. Натрий, который входит в клетку и выходит из нее, рециркулирует за счет Nа+,К+-АТФазы. Таким образом, поддерживается высокий градиент Na+ вне клетки. Транспортный белок в этих клетках сходен с белком апикальной мембраны клеток почечных канальцев, переносящих натрий. Фармакологический эффект некоторых диуретиков (фуросемида, буметанида) основан на их влиянии именно на этот белок.
Ионы хлора, которые проникают в клетку через базолатеральные участки мембраны, выходят из клетки через селективные Cl–-каналы в апикальных участках мембраны. Хлор выходит из клетки пассивно. Концентрация Cl– в клетке постоянна и составляет около 30 ммоль/л, в просвете кишки она в три раза меньше, при этом разность потенциалов — около 50-60 мВ. Поэтому при открытии каналов Cl– выходит из клетки (рис. 5-6). Выход Na+ требуемый для компенсации выхода Cl–, происходит преимущественно интерцеллюлярно под действием отрицательного электрического потенциала в просвете кишки. Надо сказать, что активная абсорбция Na+ и активная секреция Cl–, несмотря на разнонаправленный транспорт, создают отрицательный потенциал в просвете кишки (см. подпись к Рис. 5-6). Интересно отметить, что основные механизмы этой секреции встречаются у некоторых позвоночных, но не у млекопитающих, например у акул и птиц, которые имеют так называемые "солевые железы", секретирующие соль.
|
|
|
|
|
|
