 |


Гормональная регуляция деятельности мужской половой системы
|
|
|
|
Глава 20. ПОЛОВАЯ СИСТЕМА
Половая система объединяет органы, которые обеспечивают воспроизводство позвоночных и человека, и включает гонады, где происходит образование половых клеток и синтез половых гормонов, и добавочные органы полового тракта.
В мужском и женском организмах органы половой системы имеют выраженные морфофункциональные особенности, определяющие вторичные половые признаки. В мужском организме гонады представлены яичками, а добавочные органы - семявыносящими путями, семенными пузырьками, предстательной и бульбо-уретральными железами и половым членом. В женском организме гонады представлены яичниками, а добавочные органы - маткой, маточньми трубами (яйцеводами), влагалищем, наружными половыми органами. В женском организме с половым созреванием тесно связана гистофизиология молочной железы (см. главу 18).
Различия между полами предопределяются генетически через половые хромосомы (XY у мужчин и XX у женщин). Существенной особенностью женской половой системы является цикличность и периодичность деятельности. При этом созревание женской половой клетки и изменения активности секреции женских половых гормонов регулярно повторяются, тогда как мужская половая система функционирует непрерывно с момента достижения организмом половой зрелости до начала возрастного увядания.
Развитие. Закладка половой системы в начальных стадиях эмбриогенеза протекает у обоих полов одинаково (индифферентная стадия) и во взаимодействии с развитием выделительной системы (рис. 20.1). Гонада становится заметна у 4-недельного зародыша в виде половых валиков - утолщений цело-мического эпителия на вентромедиальной поверхности обеих первичных почек (mesonephros). Первичные половые клетки у зародышей обоего пола - гоноци-ты - появляются в пресомитных стадиях эмбриогенеза (во 2-й фазе гастру-ляции). Однако клетки четко выявляются при формировании желточного пузырька. В стенке последнего гоноциты характеризуются большими размерами, крупным ядром, повышенным содержанием гликогена и высокой активностью щелочной фосфатазы в цитоплазме. Здесь клетки размножаются, затем,
|
|
|
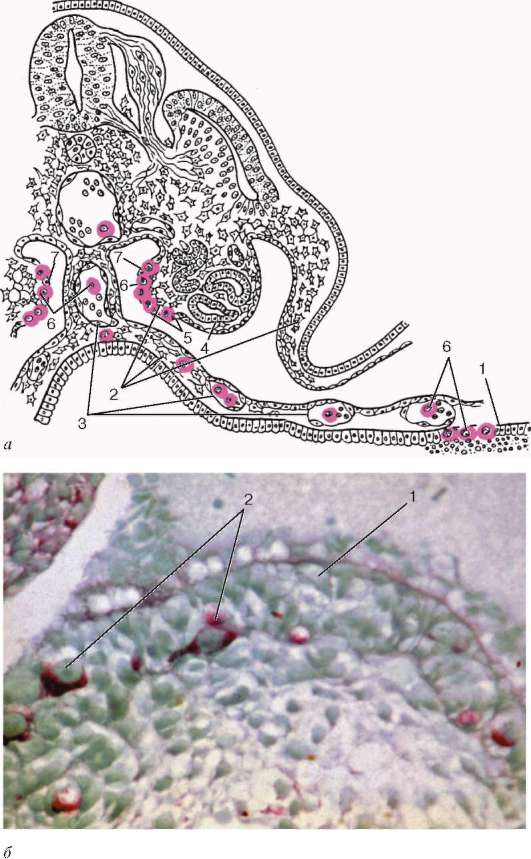
Рис. 20.1. Развитие гонад в эмбриогенезе:
а - схема первичной локализации гоноцитов (окрашены) в желточном мешке зародыша и их последующей миграции в зачаток гонад (по Пэттену, с изменениями А. Г. Кнорре): 1 - эпителий желточного пузырька; 2 - мезенхима; 3 - сосуды; 4 - первичная почка (мезонефрос); 5 - зачаток гонады; 6 - первичные половые клетки; 7 - поверхностный эпителий; б - половой валик зародыша человека 31-32 сут развития (препарат В. Г. Кожухаря): 1 - эпителий полового валика; 2 - гоноциты
продолжая деление, мигрируют по мезенхиме желточного пузырька, задней кишки и с кровотоком в толщу половых валиков. С 33-35 сут из клеток цело-мического эпителия формируются половые тяжи, которые врастают в подлежащую мезенхиму. Тяжи в своем составе содержат гоноциты. Объем гонад увеличивается, они выступают в целомическую полость, обособляются, но остаются связанными с первичной почкой. Клетки последней подвергаются апоптозу, однако часть клеток мезонефроса выселяется в окружающую мезенхиму и вступает в контакт с эпителиоцитами половых тяжей. На этой стадии развития происходит формирование гонадной бластемы, в составе которой находятся гоноциты, клетки целомического происхождения, клетки мезонефрального происхождения и клетки мезенхимы. До 7-й нед гонада не дифференцирована по полу и называется индифферентной.
В процессе развития индифферентной гонады из мезонефрального протока первичной почки, тянущегося от ее тела к клоаке, отщепляется параллельно идущий парамезонефральный проток.
|
|
|
Половые различия в строении индифферентной гонады регистрируются на 6-7-й нед эмбриогенеза человека, причем мужская гонада развивается раньше женской. Среди факторов дифференцировки мужских гонад большую роль играет Y-хромосома, на коротком плече которой локализуется ген половой детерминации (ГПД) и ряд других генов-участников детерминации пола. Экспрессия последних влияет на развитие из клеток целомическо-го происхождения поддерживающих эпителиоцитов (сустентоцитов, клеток Сертоли). Клетки Сертоли, в свою очередь, оказывают влияние на диффе-ренцировку интерстициальных эндокриноцитов (клеток Лейдига). Эти клетки обнаруживаются между половыми тяжами. Эмбриональные источники развития клеток точно не выявлены. К вероятным источникам относят клетки мезонефроса или клетки нейрального происхождения.
Начало выработки клетками Лейдига гормона тестостерона вызывает преобразование мезонефральных протоков в систему мужских половых протоков (выносящие канальцы яичка, проток придатка, семявыносящий проток, семенные пузырьки, семяизвергающий проток). В свою очередь, выработка клетками Сертоли гормона регрессии парамезонефрального протока вызывает апоптоз клеток парамезонефрального протока. На 3-м мес внутриутробного развития в срезах яичек отчетливо видны извитые тяжи, в составе которых гоноциты дифференцируются в сперматогонии.
20.1. МУЖСКАЯ ПОЛОВАЯ СИСТЕМА 20.1.1. Яички
Яички, или семенники (testes), - мужские гонады, в которых образуются мужские половые клетки и мужской половой гормон - тестостерон.
Развитие. При развитии яичка по верхнему краю первичной почки формируется будущая соединительнотканная капсула семенника - белочная
оболочка (tunica albuginea), которая отделяет половые тяжи от полового валика, давшего им начало. В дальнейшем половые тяжи развиваются в семенные канальцы (tubuli seminiferi). Семенные канальцы сливаются с канальцами семявыносящей системы, формирующейся путем перестройки эпителиальной выстилки канальцев мезонефроса. Так, канальцы сети (rete testis), приближаясь к белочной оболочке средостения, сливаются в выносящие канальцы (ductuli efferentes). Выносящие канальцы яичка, собираясь, переходят далее в канал придатка яичка (ductus epididymis), проксимальный отдел которого, многократно извиваясь, образует придаток семенника (epididymis), тогда как его дистальная часть становится семявыносящим протоком (ductus defferes). Парамезонефральный проток в мужском организме атрофируется и сохраняется только краниальный конец (формирует гидатиды, которые прикрепляются к соединительнотканной структуре яичка) и дистальный, превращающийся в мужскую маточку (utriculus prostaticus). Последняя у взрослого мужчины располагается в толще предстательной железы (рис. 20.2).
|
|
|
К концу 3-го мес завершается миграция яичек в малый таз. Опускание яичек в мошонку происходит между 6-м и 8-м мес развития.
В онтогенезе эндокринная функция яичка устанавливается раньше, чем генеративная. Мужской половой гормон - тестостерон начинает вырабатываться у зародыша человека приблизительно с 8-10-й нед внутриутробного периода. На 3-м мес эмбриогенеза клетки Лейдига в яичке достаточно многочисленны и образуют околососудистые скопления. С 6-го мес количество клеток уменьшается и остается неизменным до 2-го мес постнаталь-ной жизни.
Строение. Снаружи большая часть семенника покрыта серозной оболочкой - брюшиной, под которой располагается плотная соединительнотканная оболочка, получившая название белочной (tunica albuginea) (рис. 20.3). На задней поверхности яичка белочная оболочка утолщается, формируя средостение (mediastinum testis), от которого в глубь железы отходят соединительнотканные перегородки (septula testis), разделяющие железу на дольки (около 250 долек), в каждой из которых находится 1-4 извитых семенных канальца (tubuli seminiferi convoluti). Каждый семенной каналец имеет диаметр от 150 до 250 мкм и длину от 30 до 70 см. Приближаясь к средостению, канальцы (300-450 в каждом семеннике) сливаются и становятся прямыми, а в толще средостения соединяются с канальцами сети семенника. Из сети выходит 10-12 выносящих канальцев (ductuli efferens), впадающих в проток придатка (ductus epididymis). В дольках яичка между петлями извитых семенных канальцев находится интерстициальная (соединительная) ткань с гемо- и лимфатическими сосудами. В составе этой ткани, кроме фибробластов, обнаруживаются макрофаги, тучные клетки, а около кровеносных капилляров (преимущественно фенестрированного типа) группами располагаются гормонсинтезирующие клетки Лейдига (интерстици-альные эндокриноциты).
|
|
|
Внутреннюю выстилку канальца образует эпителиосперматогенный слой, расположенный на базальной мембране. Собственная оболочка (tunica propria) канальца представлена базальным слоем (stratum basale), миоидным слоем (stratum myoideum) и волокнистым слоем (stratum fibrosum). Кнаружи от базальной
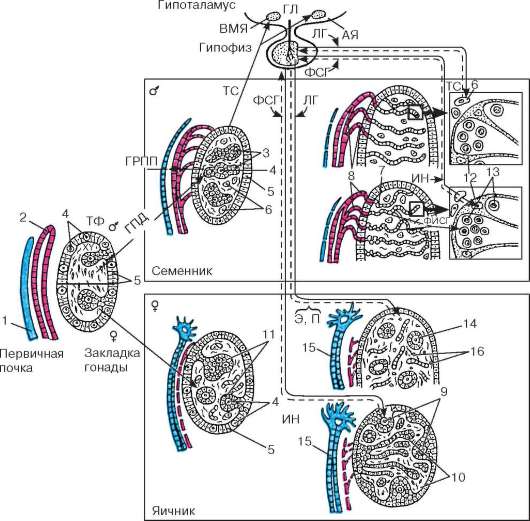
Рис. 20.2. Стадии развития гонад и становления их гормональной регуляции в онтогенезе (по Б. В. Алешину, Ю. И. Афанасьеву, О. И. Бриндаку, Н. А. Юриной): ТФ - телоферрон; ГПД - ген половой детерминации; ГРПП - гормон регрессии парамезонефрального протока; ТС - тестостерон; Э - эстрадиол; П - прогестерон; ФСГ - фолликулостимулирующий гормон; ФИСГ - фактор, ингибирующий спер-матогонии; ЛГ - лютеинизирующий гормон; ИН - ингибин; ГЛ - гонадолиберин; АЯ - аркуатное ядро; ВМЯ - вентромедиальное ядро. 1 - парамезонефральный проток; 2 - мезонефральный проток; 3 - половые тяжи; 4 - гоноциты; 5 - эпителий; 6 - клетки Лейдига; 7 - сеть семенника; 8 - выносящие канальцы семенника; 9 - корковое вещество яичника; 10 - мозговое вещество яичника; 11 - примор-диальные фолликулы; 12 - клетки Сертоли; 13 - сперматогонии; 14 - первичные фолликулы; 15 - маточная труба; 16 - интерстициальные клетки
мембраны эпителия находится сеть коллагеновых волокон базального слоя. Миоидный слой образован миоидными клетками, содержащими актино-вые филаменты. Миоидные клетки обеспечивают ритмические сокращения стенки канальцев. Наружный волокнистый слой состоит из двух частей.
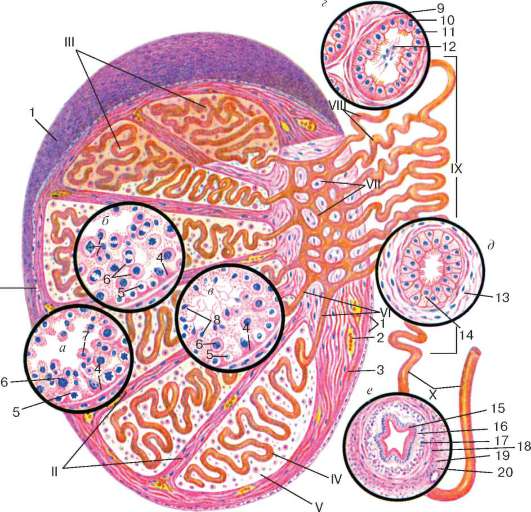
Рис. 20.3. Строение яичка (по Е. Ф. Котовскому):
а - эпителиосперматогенный слой в фазе размножения сперматогоний и в начале фазы роста сперматоцитов; б - эпителиосперматогенный слой в конце фазы роста и в фазе созревания сперматоцитов; в - фаза формирования; г - строение семявыно-сящего канальца яичка; д - строение канала придатка; е - строение семявынося-щего канала. I - оболочки яичка; II - перегородки яичка; III - дольки яичка; IV - извитой семенной каналец; V - интерстициальная ткань; VI - прямые канальцы яичка; VII - сеть яичка; VIII - выносящие канальцы яичка; IX - канал придатка; X - семявыносящий канал. 1 - мезотелий; 2 - кровеносный сосуд; 3 - соединительнотканные клетки; 4 - поддерживающие эпителиоциты (клетки Сертоли); 5 - сперматогонии; 6 - сперматоциты; 7 - сперматиды; 8 - сперматозоиды в просвете извитого семенного канальца; 9 - мышечно-волокнистая оболочка семявыносящего канальца; 10 - реснитчатые эпителиоциты; 11 - кубические эпителиоциты; 12 - сперматозоиды в семявыносящем канальце яичка; 13 - мышечно-фиброзная оболочка канала придатка яичка; 14 - двухрядный реснитчатый эпителий семявыносящего канала; 15 - двухрядный реснитчатый эпителий; 16 - собственная пластинка слизистой оболочки; 17 - внутренний продольный слой мышечной оболочки; 18 - средний циркулярный слой мышечной оболочки; 19 - наружный продольный слой мышечной оболочки; 20 - адвентициальная оболочка
|
|
|
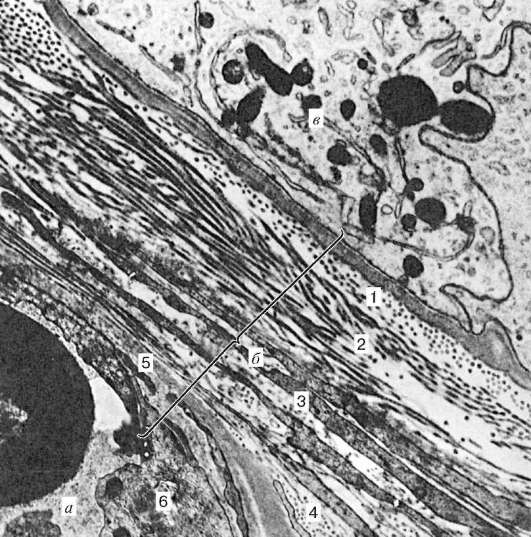
Рис. 20.4. Гематотестикулярный барьер яичка человека. Электронная микрофотография, ув. 24 000 (по А. Ф. Астраханцеву):
а - капилляр; б - гематотестикулярный барьер; в - поддерживающий эпителиоцит. 1 - базальная мембрана; 2 - внутренний волокнистый (базальный) слой; 3 - мио-идный слой; 4 - наружный волокнистый слой; 5 - базальная мембрана эндотелио-цитов; 6 - эндотелий
Непосредственно к миоидному слою примыкает неклеточный слой, образованный базальной мембраной миоидных клеток и коллагеновыми волокнами. За ними расположен слой, состоящий из фибробластоподобных клеток, прилежащий к базальной мембране эндотелиоцитов гемокапилляра.
Избирательность поступления веществ из крови в эпителиосперматоген-ный слой и различия в химическом составе плазмы крови и жидкости из семенных канальцев позволили сформулировать представление о гематоте-стикулярном барьере. Гематотестикулярным барьером называется совокупность структур, располагающихся между просветами капилляров и семенных канальцев (рис. 20.4).
Эпителиосперматогенный слой (epithelium spermatogenicum) формируется двумя клеточными дифферонами: сперматогенными клетками (cellulae spermatogenicae), которые находятся на различных стадиях дифференциров-ки (стволовые клетки, сперматогонии, сперматоциты, сперматиды и сперматозоиды) и поддерживающими эпителиоцитами (клетками Сертоли), или
сустентоцитами (epitheliocytus sustentans). Гистологические элементы двух клеточных дифферонов находятся в тесной морфофункциональной связи.
Поддерживающие эпителиоциты лежат на базальной мембране, имеют пирамидальную форму и достигают своей вершиной просвета извитого семенного канальца. Ядра клеток имеют неправильную форму с инвагинациями, ядрышко (ядрышко и две группы околоядрышкового хроматина). В цитоплазме особенно хорошо развита агранулярная эндоплазматическая сеть, комплекс Гольджи. Присутствуют также микротрубочки, микрофила-менты, лизосомы и особые кристаллоидные включения. Обнаруживаются включения липидов, углеводов, липофусцина. На боковых поверхностях сустентоциты образуют бухтообразные углубления, в которых располагаются дифференцирующиеся сперматогонии, сперматоциты и сперматиды. Между соседними поддерживающими клетками формируются зоны плотных контактов, которые подразделяют весь слой на два отдела - наружный базальный и внутренний адлюминальный. В базальном отделе расположены сперматогонии, имеющие максимальный доступ к питательным веществам, поступающим из кровеносных капилляров. В адлюминальном отделе находятся сперматоциты на стадии мейоза, а также сперматиды и сперматозоиды, которые не имеют доступа к тканевой жидкости и получают питательные вещества непосредственно от поддерживающих эпителиоцитов.
Клетки Сертоли создают микросреду, необходимую для дифференцирующихся половых клеток, изолируют формирующиеся половые клетки от токсических веществ и различных антигенов, препятствуют развитию иммунных реакций. Кроме того, они способны к фагоцитозу дегенерирующих половых клеток и последующему лизису с помощью своего лизо-сомального аппарата. Клетки синтезируют андрогенсвязывающий белок (АСБ), который транспортирует мужской половой гормон к сперматидам. Секреция АСБ усиливается под влиянием ФСГ. Поддерживающие эпителиоциты имеют поверхностные рецепторы ФСГ, а также рецепторы тестостерона и его метаболитов.
Различают два вида клеток Сертоли - светлые клетки, продуцирующие ингибин, тормозящий секрецию ФСГ аденогипофизом, и темные клетки, продуцирующие факторы, стимулирующие деление половых клеток.
Генеративная функция. Сперматогенез
Образование мужских половых клеток (сперматогенез) протекает в извитых семенных канальцах и включает четыре последовательные стадии, или фазы: размножение, рост, созревание и формирование (рис. 20.5).
Начальной фазой сперматогенеза является размножение сперматогоний, занимающих наиболее периферическое (базальное) положение в эпителио-сперматогенном слое. Среди сперматогоний выделяются два типа клеток: 1) стволовые клетки типа А; 2) клетки-предшественники типа В.
Морфологически в популяции стволовых А-сперматогоний различают светлые и темные клетки (см. рис. 20.5). Для обоих видов клеток характерно преобладание в ядрах деконденсированного хроматина и расположение ядрышек около ядерной оболочки. Однако в темных клетках типа А степень
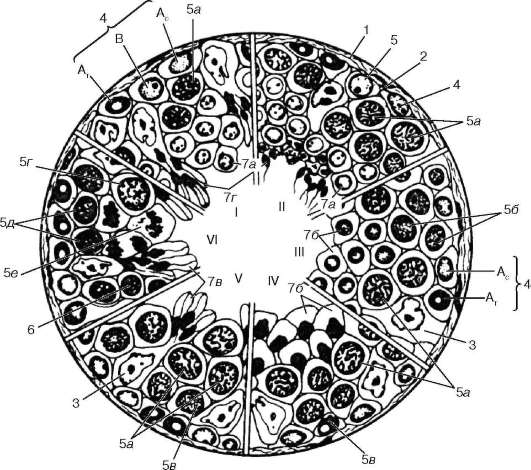
Рис. 20.5. Сперматогенез (по И. Г. Клермону, с изменениями):
I-VI - стадии цикла развития мужских половых клеток в семенных канальцах человека. 1 - соединительнотканная капсула канальца; 2 - базальная мембрана; 3 - поддерживающие клетки; 4 - сперматогонии; тип Ас - светлый; тип АТ - темный; В - тип В; 5 - сперматоциты 1-го порядка: 5а - в пахитене; 5б - в прелептотене; 5в - в лептотене; 5г - в диплотене; 5д - в зиготене; 5е - делящиеся сперматоциты 1-го порядка; 6 - сперматоциты 2-го порядка с интерфазными ядрами; 7 - сперма-тиды на различных стадиях развития (а, б, в, г)
конденсации хроматина больше, чем в светлых. Темные клетки относят к «резервным» медленно обновляющимся стволовым клеткам, а светлые - к быстро обновляющимся клеткам. Для стволовых клеток характерно наличие овальных ядер с диффузно распределенным хроматином, одного или двух ядрышек, большое содержание в цитоплазме рибосом и полисом, малое количество других органелл. Клетки типа В имеют более крупные ядра, хроматин в них не дисперсный, а собран в глыбки.
Часть стволовых клеток типа А после ряда митотических циклов становится источником развития В-сперматогоний - клеток-предшественников первичных сперматоцитов. Сперматогонии типа В после митотического деления не завершают цитокинез и остаются соединенными цитоплазмати-
ческими мостиками. Появление таких спаренных сперматогоний свидетельствует о начале процессов дифференцировки мужских половых клеток. Дальнейшее деление таких клеток приводит к образованию цепочек или групп сперматогоний, соединенных цитоплазматическими мостиками.
В следующей фазе (роста) сперматогонии перестают делиться и дифференцируются в сперматоциты 1-го порядка (первичные сперматоциты). Син-цитиальные группы сперматогоний перемещаются в адлюминальную зону эпителиосперматогенного слоя. В фазе роста сперматогонии увеличиваются в объеме и вступают в первое деление мейоза (редукционное деление). Профаза первого деления длинная и состоит из лептотены, зиготены, пахи-тены, диплотены, диакинеза.
Перед профазой в S-периоде сперматоцита 1-го порядка происходит удвоение количества ДНК. Сперматоцит находится в прелептотене. В леп-тотене хромосомы становятся видимыми в виде тонких нитей. В зиготе-не гомологичные хромосомы расположены парами (конъюгируют), образуя биваленты, между конъюгирующими хромосомами происходит обмен генами. В пахитене (от лат. pachys - толстый) пары конъюгирующих хромосом продолжают укорачиваться и одновременно утолщаться. Гомологичные хромосомы находятся в тесном контакте по всей своей длине. С помощью электронного микроскопа в сперматоцитах 1-го порядка обнаружены синаптонемные комплексы в местах соприкосновения гомологичных хромосом - парные параллельно расположенные ленты шириной около 60 нм, разделенные светлым промежутком шириной около 100 нм. В светлом промежутке видны срединная электронно-плотная линия и пересекающие его тонкие филаменты. Оба конца комплекса присоединены к ядерной оболочке. У человека формируется 23 синаптонемных комплекса. В диплотене гомологичные хромосомы, образующие бивалент, отходят друг от друга, так что становится видна каждая в отдельности, но сохраняют связь в перекрестках хромосом. Одновременно можно видеть, что каждая хромосома состоит из двух хроматид. Дальнейшая спирализация приводит к тому, что пары конъюгирующих хромосом приобретают вид коротких телец разнообразной формы - так называемых тетрад. Поскольку каждая тетрада образована двумя конъюгировавшими хромосомами, число тетрад оказывается вдвое меньше, чем исходное число хромосом, т. е. гаплоидным, - у человека 23 тетрады. В диакинезе хромосомы еще более утолщаются, после чего клетка вступает в метафазу первого деления мейоза (или первого деления созревания) и хромосомы располагаются в экваториальной плоскости. В анафазе обе хромосомы каждого бивалента расходятся к полюсам клетки - по одной к каждому полюсу. Таким образом, в каждой из двух дочерних клеток - сперматоцитов 2-го порядка (вторичных сперматоцитов) - содержится гаплоидное число хромосом (23 у человека), но каждая хромосома представлена диадой.
Второе деление созревания начинается сразу вслед за первым, и происходит как обычный митоз без репликации хромосом. В анафазе второго деления созревания диады сперматоцитов 2-го порядка разъединяются на монады, или одиночные хроматиды, расходящиеся к полюсам. В результате сперма-
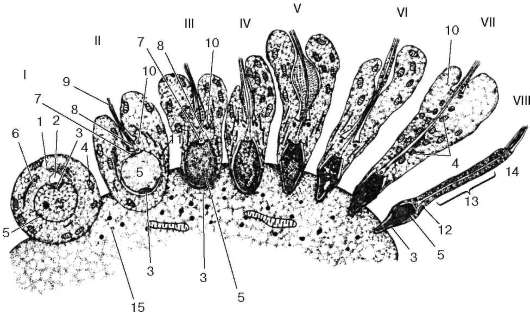
Рис. 20.6. Сперматогенез (дифференцировка сперматиды в сперматозоид) (по Б. В. Алешину):
I - сперматида, внедрившаяся в верхушку поддерживающей клетки; II-VIII - последовательные стадии формирования сперматозоида. 1 - комплекс Гольджи;
2 - акробласт; 3 - зачаток акросомы; 4 - митохондрия; 5 - ядро; 6 - центриоль; 7 - проксимальная центриоль; 8 - дистальная центриоль; 9 - трубки акронемы; 10 - кольцо; 11 - микротрубочки; 12 - шейка; 13 - митохондриальное влагалище; 14 - хвост; 15 - клетка Сертоли
тиды получают столько же монад, сколько было диад в ядрах сперматоцитов 2-го порядка, т. е. гаплоидное число. Сперматоциты 2-го порядка имеют меньшие размеры, чем сперматоциты 1-го порядка, и располагаются в средних и более поверхностных частях эпителиосперматогенного слоя.
Таким образом, каждая исходная сперматогония дает начало 4 спермати-дам с гаплоидным набором хромосом. Сперматиды больше не делятся, но путем сложной перестройки превращаются в зрелые сперматозоиды. Эта трансформация составляет суть фазы формирования (рис. 20.6).
Сперматиды представляют собой небольшие округлые клетки со сравнительно крупными ядрами. Скапливаясь около верхушек поддерживающих клеток, сперматиды частично погружаются в их цитоплазму, что создает условия для формирования сперматозоидов из сперматид. Ядро сперматиды постепенно уплотняется и уплощается.
В сперматидах около ядра располагаются комплекс Гольджи, центросома и скапливаются мелкие митохондрии. Процесс формирования сперматозоида начинается с образования в зоне комплекса Гольджи уплотненной гранулы - акробласта, прилежащего к поверхности ядра. В дальнейшем акробласт, увеличиваясь в размерах, в виде шапочки охватывает ядро, а в середине акробласта дифференцируется уплотненное тельце. Такую структуру называют акросомой. Центросома, состоящая из двух центрио-
лей, перемещается в противоположный конец сперматиды. Проксимальная центриоль прилегает к поверхности ядра, а дистальная разделяется на две части. От передней части дистальной центриоли начинает формироваться жгутик (flagellum), который затем становится осевой нитью развивающегося сперматозоида. Задняя же половина дистальной центриоли принимает вид колечка. Смещаясь вдоль жгутика, это колечко определяет заднюю границу средней, или связующей, части сперматозоида.
Цитоплазма по мере роста жгутика сползает с ядра и сосредоточивается в связующей части. Митохондрии располагаются спиралеобразно между проксимальной центриолью и колечком.
Цитоплазма сперматиды во время ее превращения в спермий сильно редуцируется. В области головки она сохраняется только в виде тонкого слоя, покрывающего акросому; небольшое количество цитоплазмы остается в области связующей части и, наконец, она очень тонким слоем покрывает жгутик. Часть цитоплазмы сбрасывается и распадается в просвете семенного канальца либо поглощается клетками Сертоли. Клетки Сертоли вырабатывают жидкость, накапливающуюся в просвете извитого семенного канальца. В эту жидкость попадают, высвобождаясь из верхушек поддерживающих клеток, сформировавшиеся сперматозоиды и вместе с ней уходят в дистальные части канальца.
Сперматогенез у человека длится около 64-75 сут и протекает на протяжении извитого семенного канальца волнообразно. Поэтому набор клеток в составе сперматогенного дифферона по ходу канальца меняется в соответствии с фазой сперматогенеза.
Реактивность и регенерация. Сперматогенез чрезвычайно чувствителен к повреждающим воздействиям. При различных интоксикациях, авитаминозах, недостаточности питания и других условиях (особенно при воздействии ионизирующим излучением) сперматогенез ослабевает и даже прекращается. Аналогичные деструктивные процессы развиваются при крипторхизме (когда семенники не опускаются в мошонку, а остаются в брюшной полости), длительном пребывании организма в среде с высокой температурой, лихорадочных состояний и особенно после перевязки или перерезки семя-выводящих каналов. Деструктивный процесс при этом поражает в первую очередь формирующиеся сперматозоиды и сперматиды. Последние набухают, нередко сливаются в характерные округлые массы - так называемые семенные шары, плавающие в просвете канальца. Так как сперматогонии и сперматоциты 1-го порядка при этом сохраняются более длительно, то восстановление сперматогенеза после прекращения действия повреждающего агента иногда оказывается возможным.
Клетки Сертоли в указанных обстоятельствах сохраняются и даже гипертрофируются, а клетки Лейдига часто увеличиваются в количестве и образуют большие скопления между запустевающими семенными канальцами.
Эндокринные функции
В рыхлой соединительной ткани между петлями извитых канальцев располагаются интерстициальные эндокриноциты (гландулоциты, клет-
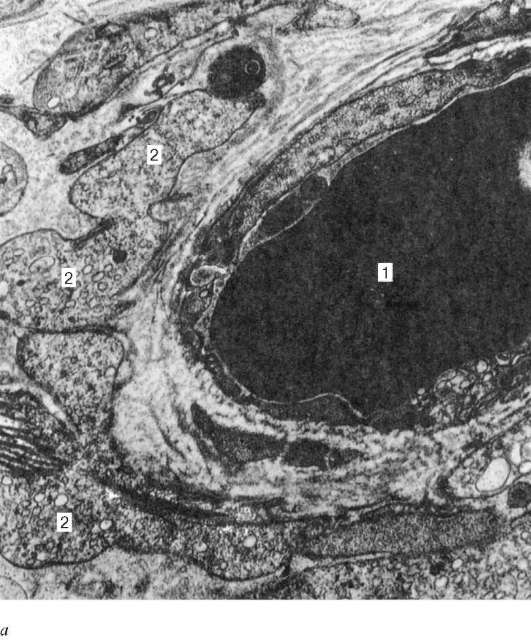
Рис. 20.7. Интерстициальные эндокриноциты (клетки Лейдига) яичка человека (по А. Ф. Астраханцеву):
а - капилляр интерстициальной соединительной ткани с прилежащими эндокри-ноцитами, увеличение 22 000; б - эндокриноцит, увеличение 10 000; в - фрагмент эндокриноцита, увеличение 26 000. 1 - капилляр; 2 - фрагменты цитоплазмы эндокриноцитов; 3 - ядро эндокриноцита; 4 - липидная капля; 5 - агранулярная эндоплазматическая сеть; 6 - строма
ки Лейдига), скапливающиеся здесь вокруг кровеносных капилляров (рис. 20.7). Эти клетки сравнительно крупные, округлой или многоугольной формы, с ацидофильной цитоплазмой, вакуолизированной по периферии, содержащей гликопротеидные включения, а также глыбки гликогена и белковые кристаллоиды в виде палочек или лент. С возрастом в цитоплазме клеток Лейдига начинает откладываться пигмент. Хорошо развитая гладкая эндоплазматическая сеть, многочисленные митохондрии с трубчатыми кри-стами указывают на способность клеток Лейдига к выработке стероидных веществ, в данном случае - мужского полового гормона.
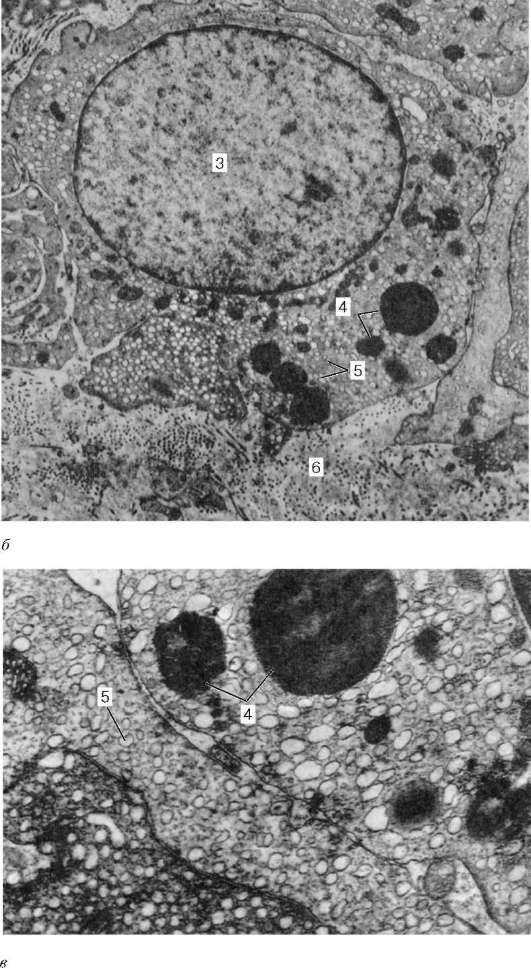
Рис. 20.7. Продолжение (обозначения см. выше)
20.1.2. Семявыносящие пути
Семявыносящие пути составляют систему канальцев (см. рис. 20.3) яичка и его придатков, по которым сперма (сперматозоиды и семенная жидкость) продвигается в мочеиспускательный канал.
Отводящие пути начинаются прямыми канальцами яичка (tubuli seminiferi recti), впадающими в сеть яичка (rete testis), располагающуюся в средостении (mediastinum). От сети отходят 12-15 извитых выносящих канальцев (ductuli effe-rentes testis), которые открываются в единый проток придатка (ductus epi-didymidis) в области головки придатка. Этот проток, многократно извиваясь, формирует тело придатка и в нижней хвостовой части становится прямым семявыносящим протоком (ductus deferens). Последний формирует ампулу семя-выносящего протока. За ампулой в проток открывается выносящий проток семенного пузырька, после чего семявыносящий проток продолжается в семя-извергающий проток. Семяизвергающий проток (ductus ejaculatorius) проникает в предстательную железу и открывается в простатическую часть мочеиспускательного канала.
Все семявыводящие пути построены по общему плану и состоят из слизистой, мышечной и адвентициальной оболочек. Эпителий, выстилающий эти канальцы, обнаруживает признаки железистой деятельности, особенно выраженной в головке придатка.
В прямых канальцах яичка эпителий образован клетками призматической формы. В канальцах сети семенника в эпителии преобладают кубические и плоские клетки. В эпителии семявыводящих канальцев чередуются группы реснитчатых эпителиоцитов с железистыми клетками, секретирую-щими по апокринному типу.
В придатке яичка эпителий протока становится двухрядным. В его составе находятся столбчатые эпителиоциты, несущие на своих апикальных верхушках стереоцилии, а между базальными частями этих клеток располагаются вставочные эпителиоциты. Эпителий протока придатка принимает участие в выработке жидкости, разбавляющей сперму во время прохождения сперматозоидов, а также в образовании гликокаликса - тонкого слоя, которым покрываются сперматозоиды. Удаление гликокаликса при эякуляции приводит к активизации сперматозоидов (капацитация). Одновременно придаток семенника оказывается резервуаром для накапливающейся спермы.
Продвижение спермы по семявыводящим путям обеспечивается сокращением мышечной оболочки, образованной циркулярным слоем гладких мышечных клеток.
Проток придатка далее переходит в семявыносящий проток (ductus deferens). Слизистая оболочка протока представлена эпителием и собственной пластинкой слизистой оболочки. Эпителий - многорядный столбчатый - включает базальные (малодифференцированные) клетки, столбчатые клетки со стереоцилиями, а также клетки, богатые митохондриями. Собственная пластинка слизистой оболочки содержит много эластических волокон. Мышечная оболочка состоит из трех слоев - внутреннего продольно-
го, среднего циркулярного и наружного продольного. В толще мышечной оболочки располагается нервное сплетение, образованное скоплением ганглиозных клеток, иннервирующих пучки гладких мышечных клеток. Сокращениями их обеспечивается эякуляция спермы. В связи со значительным развитием мышечной оболочки слизистая оболочка семявынося-щего протока собирается в продольные складки (см. рис. 20.3). Дистальный конец этого протока ампулообразно расширен. Снаружи семявыводящие пути на всем протяжении покрыты соединительнотканной адвентициаль-ной оболочкой.
Ниже места соединения семявыносящего протока и семенных пузырьков начинается семявыбрасывающий проток. Он проникает через предстательную железу и открывается в мочеиспускательный канал. В дистальной части протока эпителий становится многослойным переходным. В отличие от семявыносящего протока семявыбрасывающий проток не имеет столь выраженной мышечной оболочки. Наружная оболочка его срастается с соединительнотканной стромой предстательной железы.
Васкуляризация. Кровоснабжение семенника обеспечивается через ветвь внутренней семенной артерии, входящей в состав семенного канатика в средостение, где она разветвляется на сеть капилляров, проникающих по соединительнотканным перегородкам внутрь долек и оплетающих извитые семенные канальцы. Вокруг этих капилляров скапливаются интерстици-альные клетки.
Лимфатические капилляры также образуют сеть между канальцами семенника, а далее образуют выносящие лимфатические сосуды.
Иннервация. Нервные волокна, как симпатические, так и парасимпатические, проникают в семенник вместе с кровеносными сосудами. В паренхиме семенника рассеяны многочисленные чувствительные нервные окончания. Нервные импульсы, поступающие в семенник, способны оказывать некоторое влияние на его генеративную и эндокринную функции, но основная регуляция его деятельности осуществляется гуморальными влияниями гонадотропных гормонов аденогипофиза.
Возрастные изменения. Генеративная функция семенника начинается еще в препубертатном возрасте, но в этот период сперматогенез останавливается в начальных стадиях. Полное завершение сперматогенеза (образование сперматозоидов) наступает лишь после достижения половой зрелости - пубертатного периода. У новорожденного семенные канальцы еще имеют вид сплошных клеточных тяжей, состоящих из поддерживающих эпителиоцитов и сперматогоний. Такое строение семенные канальцы сохраняют в течение первых 4 лет постнатального периода развития мальчика. Просвет в семенных канальцах появляется лишь к 7-8 годам жизни. В это время количество сперматогоний значительно увеличивается, а к 9 годам среди них появляются одиночные сперматоциты 1-го порядка, что указывает на начало второй стадии сперматогенеза - стадии роста. Между 10 и 15 годами семенные канальцы становятся извитыми: в их просветах обнаруживаются сперматоциты 1-го и 2-го порядка и даже сперматиды, а клетки Сертоли достигают полной зрелости. К 12-14 годам заметно усиливаются
рост и развитие выводящих протоков и придатка семенника, что свидетельствует о поступлении в циркуляцию мужского полового гормона в достаточно высокой концентрации. В соответствии с этим в семенниках отмечается большое количество крупных клеток Лейдига. Возрастная инволюция семенника у мужчин происходит между 50 и 80 годами. Она проявляется в нарастающем ослаблении сперматогенеза, разрастании соединительной ткани. Однако даже в пожилом возрасте в некоторых семенных канальцах сохраняется сперматогенез и их строение остается нормальным.
Параллельно прогрессирующей атрофии эпителиосперматогенного слоя увеличивается деструкция клеток Лейдига, вследствие чего ослабевает продукция мужского полового гормона, а это в свою очередь оказывается причиной возрастной атрофии предстательной железы и частично наружных половых органов. С возрастом в цитоплазме клеток Лейдига начинает откладываться пигмент.
20.1.3. Добавочные железы мужской половой системы
К добавочным железам мужской половой системы относятся семенные пузырьки, предстательная железа, бульбоуретральные железы.
Семенные пузырьки
Семенные пузырьки - парные мешковидные структуры, развиваются как выпячивания стенки семявыносящего протока в его дистальной (верхней) части. Эти железистые органы, вырабатывающие жидкий слизистый секрет, слабощелочной реакции, богатый фруктозой, который примешивается к сперме и разжижает ее и простагландины. В стенке пузырьков имеются оболочки, границы между которыми выражены нечетко: слизистая, мышечная, адвентициальная (рис. 20.8). Слизистая оболочка собрана в многочисленные разветвленные складки, местами срастающиеся между собой, вследствие чего она приобретает ячеистый вид. Слизистая оболочка покрыта однослойным столбчатым эпителием, лежащим на тонкой базальной мембране. В составе эпителия различают столбчатые и базаль-ные эпителиоциты. В собственной пластинке слизистой оболочки много эластических волокон. В слизистой оболочке расположены терминальные отделы желез альвеолярного типа, состоящие из слизистых экзокриноцитов (exocrinocytus mucosus).
Мышечная оболочка хорошо выражена и состоит из двух слоев гладких мышечных клеток - внутреннего циркулярного и наружного продольного. Адвентициальная оболочка состоит из плотной волокнистой соединительной ткани с большим содержанием эластических волокон.
Предстательная железа
Предстательная железа, или простата (prostata), - мышечно-железистый орган, охватывающий верхнюю часть мочеиспускательного канала (уре-
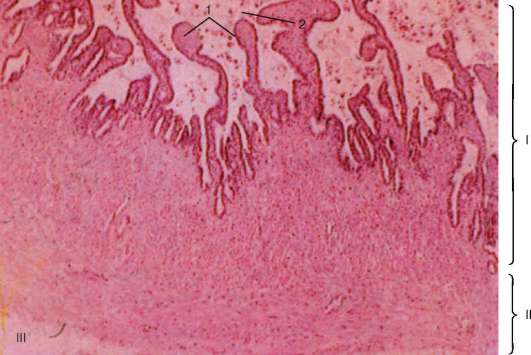
Рис. 20.8. Семенной пузырек:
I - слизистая оболочка; II - мышечная оболочка; III - наружная соединительнотканная оболочка. 1 - складки слизистой оболочки; 2 - секрет в просвете железы
тры), в которую открываются протоки многочисленных простатических желез.
Развитие. У человека формирование предстательной железы начинается на 11-12-й нед внутриутробного развития, при этом из эпителия мочеиспускательного канала в окружающую мезенхиму врастают 5-6 тяжей. В первой половине пренатального эмбриогенеза из разрастающихся эпителиальных тяжей развиваются преимущественно альвеолярно-трубчатые простатические железы. В процессе развития многослойный эпителий желез под влиянием андрогенов становится многорядным, в составе которого возникают диффероны секреторных, слизистых и эндокринных клеток. Базальные эпителиоциты являются камбиальными. Со второй половины эмбриогенеза преобладает рост гладкой мышечной ткани и соединительнотканных прослоек предстательной железы. Просветы в эпителиальных тяжах появляются в конце предплодного периода развития зарод
|
|
|
