 |


«Классификация жизненных форм растений по Раункиеру»
|
|
|
|
Вопрос №21.
«Классификация жизненных форм растений по Раункиеру»
Жизненная форма – это внешняя форма вида (организма, сообщества), приобретенная им в процессе эволюции и отражающая способ взаимодействия его со средой обитания, причем не с одним из факторов, а с их комплексом. Жизненная форма вырабатывается в ходе вековой эволюции видов и характеризирует адаптацию вида к определенному образу жизни.
Классификация жизненных форм растений К. Раункиера:
Сходные типы приспособлений растений к среде – это сходные способы перенесения наиболее трудных условий, признак – расположение почек или верхушек побегов в течение неблагоприятного времени года по отношению к поверхности почвы или снегового покрова):
- эпифиты – растут на других растениях и не имеют корней в почве;
- фанерофиты (деревья, кустарники, лианы) – оказываются зимой над поверхностью снега, их почки защищены покровными чешуйками (тополь);
- хамефиты – существуют в виде ползучих или несколько приподнятых стеблей, их почки зимой частично или полностью прикрывает снег (черника);
- хемикриптофиты – сохраняют почки у самой поверхности почвы среди высохшей прошлогодней растительности, зимой их прикрывает снег (злаки);
- криптофиты (геофиты) – наземная часть их отмирает, сохраняя почки в клубнях, луковицах, корневищах, скрытых в почве и хорошо защищенных (тюльпан);
- терофиты – однолетние растения, отмирающие с наступлением неблагоприятного сезона; выживают лишь семена и споры, прорастающие при наступлении благоприятных условий (фасоль);
- гидрофиты – все водные растения из разных систематических групп.
Вопрос №22.
|
|
|
«Концепция максимальной ёмкости среды. Внутривидовая конкуренция»
Когда численность популяции стабилизируется на определенном, или стационарном уровне, говорят, что для нее достигнут предел емкости среды.
Емкость среды – способность природного окружения обеспечивать нормальную жизнедеятельность в единице пространства определенного числа особей без заметного нарушения самого окружения. Емкость среды проявляет свое действие через лимитирующие факторы – запасы пищи, минеральных солей, пространства, влияние особей друг на друга в результате повышения плотности. Поддержание численности на определенном уровне происходит посредством механизмов отрицательной обратной связи. Емкость среды может быть определена довольно просто или может являться производным чрезвычайно сложных биотических и абиотических взаимосвязей.
Концепция ёмкости среды была предложена ещё А. Леопольдом (1933) в 30-х годах ХХ века и активно развивается в рамках рационального природопользования. При заполнении емкости среды до максимума значительная часть особей находится в состоянии уязвимости в результате любого возможного негативного воздействия, что может стать причиной резкого падения численности.
Именно поэтому, концепция максимальной емкости среды связан с другим понятием общей экологии – конкуренцией.
Конкуренция – это взаимоотношения видов со сходными экологическими требованиями существующих за счет общих ресурсов, имеющихся в недостатке. Когда такие виды обитают совместно, каждый из них находится в невыгодном положении, так как присутствие другого уменьшает возможности в овладении пищей, убежищами и прочими средствами к существованию, которыми располагает местообитание. Конкуренция – единственная форма экологических отношений, отрицательно сказывающаяся на обоих взаимодействующих партнерах.
|
|
|
Формы конкурентного взаимодействия могут быть самыми различными: от прямой физической борьбы до мирного совместного существования. Тем не менее если два вида с одинаковыми экологическими потребностями оказываются в одном сообществе, рано или поздно один конкурент вытесняет другого. Это одно из наиболее общих экологических правил, которое получило название закона конкурентного исключения и было сформулировано Г. Ф. Гаузе.
Внутривидовая конкуренция - соперничество за одни и те же ресурсы, происходящее между особями одного и того же вида. Это важный фактор саморегулирования популяции. Примером может стать конкуренция птиц одного вида из-за места гнездования. Самцы многих видов млекопитающих(например, оленей) в период размножения вступают друг с другом в борьбу за возможность обзавестись семьёй.
Двустороннее, взаимное угнетающее действие одних организмов на других имеет всегда, когда совпадают их экологические ниши и когда ограничена емкость среды. Совпадение ниш может быть абсолютным, когда речь идет об организмах одного вида, даже одной популяции, о внутривидовой конкуренции.
Вопрос №23.
«Динамика популяций. Скорость естественного роста популяции, кривые роста»
Динамика популяций – это процессы изменения ее основных биологических показателей (численности, биомассы, структуры) во времени в зависимости от экологических факторов.
Скорость роста популяции — изменение численности популяции за единицу времени. Скорость роста популяции может быть положительной, нулевой и отрицательной. Она зависит от показателей рождаемости, смертности и миграции. Если рождаемость в популяции превышает смертность, то популяция, как правило, будет расти. С увеличением плотности скорость роста популяции постепенно снижается до нуля. При нулевом росте популяция стабильна, т. е. размеры ее не меняются. Отдельные организмы при этом могут расти и размножаться. Нулевая скорость роста означает лишь то, что скорость размножения, если оно происходит, уравновешена смертностью. Данная картина характерна для ряда одноклеточных и многоклеточных организмов, например для клеток водорослей в культуральной жидкости, для фитопланктона озер и океанов весной, для насекомых.
|
|
|
Таким образом, скорость роста популяции в естественных местообитаниях будет зависеть от климатических изменений, от снабжения пищей и от того, ограничено ли размножение определенным временем года и др., что должно учитываться при составлении моделей или их усовершенствовании.
Скорость роста популяции при отсутствии каких-либо экологических ограничений описывает дифференциальное уравнение:
dN/dф = rN, (1)
где N - численность особей в популяции; ф - время; r - константа скорости естественного прироста.
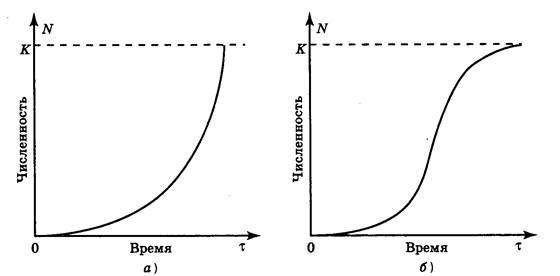
J-образная модель роста популяции. Если r > 0, то со временем численность популяции становится больше. Рост происходит сначала медленно, а затем стремительно увеличивается по экспоненциальному закону, т. е. кривая роста популяции принимает J-образный вид (рис. 1, а).
S-образная модель роста популяции. Иное развитие получает ситуация при ограниченности пищевых ресурсов либо при скоплении токсичных продуктов (отходов) метаболизма. Первоначальный экспоненциальный рост в исходных благоприятных условиях со временем продолжаться не может и постепенно замедляется. Плотность популяции регулирует истощение пищевых ресурсов, накопление токсикантов и поэтому влияет на рост численности. С увеличением плотности скорость роста популяции постепенно снижается до нуля, и кривая выходит на некоторый стабильный уровень (график образует плато). Кривая такого роста (рис. 1, б) имеет S-образную форму, и поэтому соответствующая модель развития событий называется S-образной.
Скорость роста численности в S-образной модели определяет дифференциальное уравнение:
dN/dф = rN(l - N/K), (2)
где К - поддерживающая емкость среды, т. е. максимальный размер популяции, которая может существовать в данных условиях, удовлетворяя свои потребности неопределенно долго.
Если N > К, скорость роста отрицательна. Если N < К, скорость роста положительна и величина популяции N стремится к К, т. е. приводится в соответствие с поддерживающей емкостью среды. Если N = К, скорость роста популяции равна нулю. При нулевом росте популяция стабильна, т. е. ее размеры не меняются, хотя отдельные организмы по-прежнему растут, размножаются и отмирают. Происходящее размножение уравновешивается смертностью.
В специализированной литературе J- и S-образные модели роста численности часто называют соответственно экспоненциальной и логистической.
|
|
|
