 |


Перекрестные эффекты адаптации к стрессорным ситуациям
|
|
|
|
Адаптация к стрессорным воздействиям вызывает увеличение мощности ряда стресс-лимитирующих систем центрального и периферического действия. Метаболиты этих систем от опиоидных пептидов и ГАМК до простагландинов и естественных антиоксидантов обладают широким спектром адаптивных и защитных эффектов; это становится основой положительной перекрестной адаптации организма к самым различным факторам окружающей среды, которая защищает его от новых неиспытанных стрессорных ситуаций, ишемических повреждений, которые обычно сопровождаются стрессом, и даже от прямых химических повреждений клеток и тканей. Так, показано, что адаптация к повторным иммобилизационным стрессорным воздействиям защищает от качественно иной стрессорной ситуации — от так называемого «насильственного плавания», которое состоит в том, что мыши или крысы вынуждены плавать в цилиндре, из которого нельзя
уйти; некоторое время спустя животные занимают так называемую иммобильную позицию, т. е. прекращают попытки выбраться из воды посредством активного плавания и начинают держаться на воде пассивно, по-видимому, за счет воздушной подушки в волосяном покрове. Время до возникновения «реакции иммобильности» пропорционально устойчивости к стрессорному воздействию; оно увеличивается под влиянием антидепрессантов и служит показателем их эффективности [Platt J. E., Stone E. А., 1982].
Предварительная адаптация к коротким иммобилизационным стрессорным воздействиям увеличивает «время иммобильности» так же, как антидепрессанты. Причем в основе защитного эффекта обоих факторов лежит одно и то же явление — десенситизация, реализовавшаяся в различных областях мозга крыс и проявлявшаяся уменьшением там плотности β-рецепторов [Stone E., Platt J., 1982 а] и соответственно снижением активности норадреналинзависимой аденилатциклазы [Stone E., Platt J., 1982 b]. Эти изменения надо рассматривать как результат действия метаболитов стресс-лимитирующих систем или глубокой перестройки в липидном бислое нейрональных мембран.
|
|
|
Адаптация к коротким стрессорным воздействиям, как было показано, закономерно приводит к увеличению физиологической мощности симпатико-адреналовой регуляции. Поскольку мобилизация этой регуляторной системы составляет первое и необходимое звено адаптации к основным факторам среды, например к физическим нагрузкам, холоду, гипоксии, можно предположить, что адаптация к стрессорным ситуациям повышает резистентность ко всем перечисленным факторам, т. е. обладает положительным перекрестным действием. Работы A. Kuroshima и соавт. (1984) свидетельствуют, что это положение верно по крайней мере для адаптации к холоду. Исследователи исходили из предположения, что грызуны, как и человек, адаптируются к холоду за счет увеличенной термогенетической активности, не связанной с мышечными сокращениями, т. е. за счет термогенеза без дрожи. Такая метаболическая стратегия в основном находится под контролем катехоламинов. Однако другие гуморальные факторы, такие как тиреоидные, адренокортикальные и панкреатические гормоны, возможно принимают участие в формировании и поддержании термогенеза без дрожи при холодовой адаптации [Jansky L., 1973; Kuroshima A. et al., 1978]. Причем важным фактором этой адаптивной реакции у млекопитающих является значительный гормонально-индуцированный рост бурой жировой ткани, локализованной в межлопаточном пространстве [Smith R. Е., 1961], которая метаболизируется и служит источником тепловой энергии во время действия холода.
Эксперименты A. Kuroshima и соавт. заключались в том, что взрослых крыс-самцов линии Впстар адаптировали к иммобилизационному стрессу путем привязывания их к доске на 2 ч в день 8 дней в нед в течение 1—8 нед. Адаптированные крысы меньше набирали массу в течение эксперимента по сравнению с контролем, но у них наблюдали увеличение резистентности к холоду. Это выражалось в отсутствии достоверного снижения температуры в ободочной кишке во время пребывания животных при —5 ° Св
|
|
|
течение 5 ч, тогда как температура в ободочной кишке контрольных животных в таких условиях прогрессивно падала. У адаптированных крыс был достоверно потенцирован термогенез без дрожи, что оценивалось по величине вызванного норадреналином изменения потребления кислорода. У этих животных масса и содержание белка в межлопаточной бурой жировой ткани увеличивались, и митохондрии в этой ткани были плотнее «упакованы». Удаление межлопаточной бурой жировой ткани приводило к значительному снижению резистентности к холоду, так как уменьшало термогенез?ез дрожи.
Таким образом, несомненно, что адаптация к стрессорным воздействиям вызывает гормонально-индуцированный рост органа термогенеза — бурой жировой ткани и тем самым повышает резистентность к холоду. В итоге реализуется феномен положительной перекрестной адаптации. По-видимому, за счет аналогичного механизма, т. е. за счет повышения физиологической, мощности стресс-реализующих систем, адаптация к повторным стрессорным воздействиям повышает устойчивость сердца к максимальной нагрузке.
В работе, посвященной изучению этого вопроса, мы совместно с Л. М. Белкиной изучали сократительную функцию сердца крыс в условиях острого опыта in situ путем электроманометрической регистрации давления в левом желудочке.
На первом этапе эксперимента регистрировали сократительную функцию сердца в условиях относительного физиологического покоя. На втором этапе оценивали реакцию сердца на максимальную изометрическую нагрузку, вызванную пережатием восходящей аорты на 30 с. Используя пережатие аорты как прием для создания максимальной нагрузки на сердце, следует иметь в виду, что мобилизация функции сердца при этом обусловлена не только внезапно возросшим сопротивлением изгнанию и включением механизмов саморегулирования, но также возбуждением адренергической системы, обусловленным ишемией мозга.
|
|
|
Данные, представленные на рис. 12, отражают основные результаты этих исследований ипозволяют отметить два положения. Первое из них заключается в том, что адаптация к коротким иммобилизационным стрессорным воздействиям не вызывала достоверных изменений сократительной функции левого желудочка сердца в условиях относительного физиологического покоя. Отмечалась лишь некоторая тенденция к снижению развиваемого давления и скорости расслабления. Второе — основное — положение состоит в том, что адаптация к коротким стрессорным воздействиям существенно повысила резистентность сердца к максимальной нагрузке, вызванной пережатием аорты. Это явление выражалось в том, что на 5-й секунде пережатия все определявшиеся параметры сократительной функции были практически одинаковы в контроле и у адаптированных животных, а на 25-й секунде пережатия, когда в контроле развивалась выраженная депрессия сократительной функции, у адаптированных животных эта депрессия была незначительной. В результате развиваемое систолическое давление у адаптированных животных оказалось увеличен-
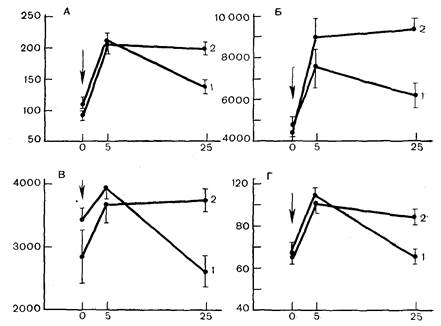
|
| Рис. 12. Показатели сократительной функции левого желудочка у крыс при пережатии аорты. А — развиваемое систолическое давление, мм рт. ст.; Б — максимальная скорость развитая давления в систолу мм рт. ст. /с; В — максимальная скорость снижения давления в диастолу мм рт. ст. /с; Г — ИФС мм рт. ст..мг— 1.мин-1. Ординаты — величины показателей (А — Г); абсциссы — время с момента пережатия аорты, с (стрелка). 1 — контроль; 2 — адаптация. |
ным на 43%, ИФС1 — 43%, скорость сокращения почти на 80%. а скорость расслабления на 77% по сравнению с контролем.
Важно, что такое сохранение более высокого уровня сократительной функции сердца у адаптированных к стрессу животных реализовалось в условиях сниженного почти на 40% диастолического давления, и следовательно, едва ли может быть обусловлено более выраженным включением таких механизмов саморегуляции сердца, как механизм Старлинга.
|
|
|
При объяснении обнаруженного факта надо иметь в виду, чтоадаптация к повторным стрессорным воздействиям закономерновлечет за собой стационарную активацию синтеза катехоламинов в мозгу и надпочечниках, т. е. в конечном счете приводит к увеличению мощности симпатико-адреналовой системы. В соответствии с этим можно предположить, что одним из важных факторов, обеспечивающих сохранение высокой функции сердца при длительной максимальной нагрузке у адаптированных к стрессу
1 ИФС — интенсивность функционирования структур сердца, отражающая величину сократительной функции сердца, отнесенную к ед. массы.
животных, является более стойкое, чем в контроле, положительное инотропное влияние на сердце адренергической системы.
Таким образом, сердце животных, адаптированных к коротким стрессорным воздействиям, обладает существенно повышенной устойчивостью к максимальным нагрузкам, и явление это с большой долей вероятности может быть объяснено увеличенной мощностью адренергического регуляторного аппарата сердца и систем, которые обеспечивают его экономное функционирование.
Другой практически достаточно актуальный пример положительной перекрестной резистентности, а именно возможность ограничения нарушений сократительной функции сердца при экспериментальном инфаркте с помощью предварительной адаптации к повторным стрессорным воздействиям, был также изучен в нашей лаборатории. Основой этих исследований явилось развитие раннего представления о роли стресс-реакции в патогенезе ишемической болезни сердца [Меерсон Ф. 3., Пшенникова М. Г., Уголев A. A., 1982J, в соответствии с которым стресс-реакция может включаться в патогенетическую цепь ишемической болезни первично — под влиянием внешних факторов, а именно эмоциональных ситуаций окружающей среды, и вторично, под влиянием таких эндогенных факторов, как боль и страх смерти, вызванных коронарным приступом. Возникшая стресс-реакция может потенцировать развитие коронарного приступа или инфаркта миокарда многими путями. Так, известно, что тяжелый стресс активирует развитие атеросклероза, вызывает некоронарогенное повреждение миокарда, может привести к контрактурному спазму коронарных артерий, играет роль в увеличении свертываемости крови и развитии коронаротромбоза, снижает резистентность миокарда к гипоксии, вызывает острую перегрузку сердца за счет роста периферического сопротивления, приводит к патологическому депонированию крови за счет снижения тонуса емкостных сосудов и депрессии сократительной функции воротной вены. Далее стресс вызывает гипервентиляцию, а связанный с нею алкалоз приводит к доказанному теперь дополнительному сужению коронарных артерий. Наконец, хорошо известен аритмогенный эффект стресса, его роль в возникновении остановки сердца. Это многообразное включение стресс-реакции в патогенетическую цепь инфаркта миокарда делало вероятным, что адаптация к стрессорным ситуациям может увеличить резистентность сердца к самому инфаркту.
|
|
|
Результаты экспериментов [Меерсон Ф. 3., Белкина Л. М., Заяц В. И., 1985] свидетельствуют, что адаптация к коротким иммобилизационым стрессорным воздействиям существенным образом уменьшает нарушения сократительной функции сердца, выявляющиеся после создания экспериментального инфаркта миокарда как в условиях относительного физиологического покоя, так и при максимальной нагрузке на сердце. Изучение сократительной функции сердца проводилось в условиях острого опыта in situ под уретановым наркозом путем электроманометрической регистрации давления в полости левого желудочка. Выяснилось,
что под влиянием инфаркта миокарда, воспроизведенного по Г. Селье и соавт. (1961), в условиях покоя наблюдается снижение на 25—30% таких важных показателей функции левого желудочка, как развиваемое давление, интенсивность функционирования структур (ИФС), скорости развития и снижения давления. У животных, предварительно адаптированных к коротким стрессорным воздействиям, депрессия этих параметров сократительной функции при инфаркте оказалась недостоверной. Такого рода эффект сохранялся и при максимальной нагрузке на сердце, которую создавали полным пережатием аорты на 30 с. Как и в описанной выше работе, было показано, что сама по себе адаптация к коротким стрессорным воздействиям оказалась фактором, существенно повысившим сократительную функцию левого желудочка в условиях такой максимальной нагрузки. При максимальной нагрузке на левый желудочек с инфактом этот эффект предварительной адаптации оказался еще более значительным. Действительно, по сравнению с неадаптированными животными, имевшими инфаркт, у адаптированных животных, которым также воспроизвели инфаркт, развиваемое левым желудочком давление на 25-й секунде пережатия аорты было увеличено в 2,8 раза (р<0,001), ИФС в 3,8 раза (р<0,01), максимальные скорости развития и снижения давления в 3,7 и 3,25 раза соответственно (р<0,001) (рис. 13).
Таким образом, несомненно, что предварительная адаптация к коротким стрессорным воздействиям резко ограничивает нарушения сократительной функции левого желудочка сердца при экспериментальном инфаркте и является эффективным фактором защиты от ишемического повреждения. Этот защитный эффект может быть обусловлен как уменьшением зоны некроза, так и предупреждением повреждения, которое обычно развивается при инфаркте в неишемизированных отделах сердца.
Для того чтобы оценить возможность защиты неишемизированных отделов миокарда при инфаркте у адаптированных животных, было детально исследовано влияние предварительной адаптации к коротким стрессорным воздействиям на функцию изолированного правого предсердия животных, у которых был воспроизведен инфаркт левого желудочка. При этом совершенно определенно выявляется вызываемая инфарктом депрессия показателей сократительной функции правого предсердия. Предварительная адаптация к коротким стрессорным воздействиям сама по себе достоверно не влияла на сократительную функцию миокарда правого предсердия, но в значительной мере предотвращала депрессию его сократительной функции при инфаркте левого желудочка. Действительно, у неадаптированных животных инфаркт приводил к снижению максимальной развиваемой предсердием силы и ИФС примерно в 2—2,5 раза, а у адаптированных животных это снижение составляло лишь менее 1/3 от контрольного уровня. Таким образом, несомненно, что предварительная адаптация к коротким стрессорным ситуациям ограничивала

|
| Рис. 13. Показатели сократительной функции левого желудочка у крыс с инфарктом миокарда при пережатии аорты. А — развиваемое систолическое давление мм рт. ст.; Б — максимальная скорость развития давления в систолу мм рт. ст. /с; В — максимальная скорость снижения давления в диастолу, мм рт. ст. /с; Г — ИФС мм рт. ст..мг-1.мин-1. Ординаты — величины показателей (А— Г), абсциссы — время с момента пережатия аорты, с (стрелка). 1 — контроль; 2 — инфаркт; 3 — а даптация-i- инфаркт. Заштрихованная зона — защитный эффект адаптации. |
нарушение функции неишемизированных отделов миокарда и сердца в целом при экспериментальном инфаркте миокарда.
Имеются данные о том, что это явление представляет собой не просто экспериментальный феномен. В эпидемиологическом исследовании L. Wilhelmsson и соавт. (1982) было показано, что у людей, существование которых до инфаркта миокарда было богато стрессорными ситуациями, длительность жизни после инфаркта была достоверно больше, чем у людей с относительно спокойным образом жизни. Аналогичные данные ранее были представлены С. Jenkins и соавт. (1976), которые показали, что люди, обладающие повышенной эмоциональной реакцией на возникающие жизненные ситуации, имеют меньшую смертность при первом и повторных инфарктах миокарда, чем люди с умеренной реакцией. Разумеется, эти клинические наблюдения требуют проверки и развития, однако, факт, что высокий уровень адаптации к стрессорным ситуациям, по-видимому, повышает резистентность к вероятному инфаркту миокарда, заслуживает внимания.
При оценке приведенных выше экспериментальных данных и эпидемиологических наблюдений, свидетельствующих о защитном эффекте предварительной адаптации к стрессорным ситуациям при инфаркте миокарда, следует подчеркнуть, что решающая роль
в механизме этого эффекта несомненно принадлежит предупреждению и ограничению сопутствующей инфаркту стресс-реакции и вызываемых ею повреждений. Это кардиопротекторное действие адаптации обеспечивается за счет компонентов ее структурного «следа», приводящих, как было указано выше, к повышению функциональных возможностей стресс-лимитирующих систем организма. Главное место в ограничении стресс-реакции занимают ГАМК-ергическая, серотонинергическая система и система опиоидных пептидов. Повышение активности именно этих систем обеспечивает адаптированному организму при инфаркте миокарда уменьшение степени повреждающего действия катехоламинов и, кроме того, обеспечивает более выраженный анальгетический эффект, что также способствует ограничению стресс-реакции.
Существенную роль в ограничении стрессорных повреждений неишемизированных отделов сердца и самих ишемических его повреждений при инфаркте в адаптированном организме играет повышение мощности антиоксидантных систем. Именно за счет этого структурного «следа» адаптации в нем ограничивается активация свободно-радикального окисления, играющая решающую роль в патогенезе стрессорных, ишемических и реоксигенационных повреждений миокарда при инфаркте. Более подробно этот аспект защитного действия адаптации будет рассмотрен в 6-й главе. Можно полагать, что в защитном эффекте адаптации к стрессорным воздействиям участвуют также и другие стресс-лимитирующие системы, в частности система простагландинов, ограничивающая тромбообразование, и т. д.
Наиболее яркий пример перекрестной резистентности при адаптации к стрессу, по-видимому, составляет адаптивное повышение резистентности слизистой желудка у адаптированных животных к прямому повреждающему действию химических факторов. В исследованиях J. Wallace и М. Cohen (1984), показано, что адаптация к мягкому длительному стрессу, обусловленная минимальной иммобилизацией, обладает таким защитным эффектом. Для создания минимальной иммобилизации в заднюю часть шеи крыс под наркозом подкожно вживляли металлическое кольцо диаметром 2 см. Проволока длиной 25 см соединяла это кольцо с потолком клетки. Крысы имели свободный доступ к пище и могли двигаться по клетке. В контрольной группе была сделана ложная операция. Через 2, 5 и 10 дней крысам вновь давали наркоз и из открытого желудка делали камеры ex vivo. Слизистую желудка последовательно помещали в растворы маннитола, этанола и НС1, изотоничный маннитолу.
Были подобраны такие концентрации этанола и НС1, которые в контроле повреждают 100% поверхности слизистой желудка. В конце каждого эксперимента слизистую фотографировали и измеряли площадь повреждений планиметрически (% площади слизистой с повреждениями). У всех адаптированных животных повреждение было существенно меньше, чем в контроле. Защитный эффект возрастал с увеличением длительности адаптации: два дня адаптации уменьшали повреждения на 14%; 5 дней — на 49%, а 10 дней — на 95%. Предварительное введение крысам индометацина (5 мг/кг внутрибрюшинно) полностью устраняло защитный эффект адаптации.
Таким образом, данный защитный эффект зависел от адаптивного увеличения мощности или эффективности системы проста-
гландинов. Действительно, дальнейшие исследования показали, что сразу же после возникновения минимальной иммобилизации: в слизистой дна желудка возникает активация биосинтеза простагландинов группы Е. Через 7 сут после начала адаптации уровень синтеза оказывается увеличенным в 3 раза и выходит на плато. Таким образом, эти исследования показали, что цитопротекторный эффект адаптации к стрессорным воздействиям достигается за счет адаптивного увеличения мощности конкретной стресслимитирующей системы местного действия, а именно системы простагландинов.
Другой, по-видимому, перспективный перекрестный защитный эффект адаптации к стрессорным воздействиям был впервые установлен в исследованиях Ю. И. Зимина (1970) и И. А. Рудакова (1963), которые показали, что при повторных стрессорных воздействиях, т. е. при ситуации, которую обозначали как хронический стресс, повышена резистентность к ионизирующей радиации. В дальнейшем G. Mickley н соавт. (1983) показали, что повторные стрессорные воздействия увеличивают толерантность к ионизирующим излучениям, а именно уменьшают налоксончувствптельную двигательную гиперактивность, обнаруженную после облучения. Можно полагать, что этот перекрестный защитный эффект получит в будущем развитие в области радиобиологии.
Вместе с тем адаптация к стрессорным ситуациям, сопряженная с увеличенным распадом структур и имеющая заведомо значительную структурную «цену», далеко не всегда приводит к положительным перекрестным эффектам. Известно, что повторные стрессорные воздействия, даже при успешной адаптации к ним, могут приводить к снижению содержания тестостерона в плазме крови у разного вида животных [Rose В. et al., 1972; Repcekova D., Mikulay L., 1977; Gray G. et al., 1978], в том числе у человека [Kreuz L. et al., 1972; Davidson J. et al., 1978]. В более поздних исследованиях G. Charpent и соавт. (1981) показано, что иммобилизационный стресс, воспроизводимый в течение 10 дней по 6 ч в день вызывал у крыс значительное снижение содержания тестостерона не только в плазме крови, но и в семенниках, причем это изменение, по-видимому, было обусловлено глубокими изменениями в клетках самих семенников, так как в условиях in vitro в препаратах, обогащенных интерстициальными клетками адаптированных животных, базальный уровень продукции тестостерона, а также увеличение его секреции в ответ на гонадотропин и бутирил-цАМФ были резко снижены. Этим, по-видимому, и объясняется установленный далее в этой работе факт, что у контрольных животных увеличение в плазме крови содержания тестостерона в ответ на введение гонадотропина возникало при дозе этого препарата, равной 5 международным единицам, а у животных, адаптированных к стрессу указанным способом, только при дозе 10 и 50 таких единиц.
Авторы пришли к заключению, что повторные иммобилизационные стрессы снижают чувствительность гранулоцитов яичка к гонадотропину.
Столь значительная «цена» адаптации к стрессорным воздействиям, которая по существу ставит под вопрос продолжение рода, на наш взгляд, реализуется лишь при чрезмерной интенсивности и длительности таких воздействий. Это явление, так же как и бесспорный факт существования опасного семейства стрессорных заболеваний, само по себе не исключает возможности использования адаптации к стрессорным воздействиям и ее положительных перекрестных эффектов. Вместе с тем очевидно, что явление «цены» адаптации к стрессу выдвигает на первый план вопрос о пра-
вильном дозировании и выборе формы и длительности стрессорных воздействий в процессе медицинских мероприятий или профессиональной подготовки.
В целом, рассмотренные в этой главе стресс-лимитирующие системы, связанные с их активацией перекрестные эффекты адаптации и, наконец, защитные эффекты метаболитов этих систем явились основанием для изучения возможности использования адаптации к коротким стрессорным воздействиям и указанных метаболитов для предупреждения и устранения наиболее опасных нарушений сердечной деятельности, возникающих под влиянием ишемии и стресса, — для предупреждения аритмий и фибрилляции сердца.
Результаты этих исследований представлены в последующих главах.
ГЛАВА 5.
ПРЕДУПРЕЖДЕНИЕ ФИБРИЛЛЯЦИИ СЕРДЦА ПРИ ПОМОЩИ АДАПТАЦИИ К СТРЕССОРНЫМ СИТУАЦИЯМ И ДРУГИМ ФАКТОРАМ СРЕДЫ
Ишемию миокарда обычно определяют как состояние, при котором потребность сердца в кислороде превышает его поступление к кардиомиоцитам. В свете современных представлений о регуляции сердца, сосудов и системы крови это определение само по себе подразумевает важную роль нарушений нейроэндокринной регуляции в возникновении ишемической болезни. Действительно, атеросклероз коронарных сосудов и множественные их стенозы являются результатом генетически и алиментарно детерминированных нарушений регуляции обмена холестерина. Спазм коронарных артерий, быстро и нередко значительно ограничивающий их пропускную способность, составляет результат сильных адренергических воздействий через α-рецепторы или нарушения регуляторных вазодилататорных влияний аденозина и простагландинов.
Аналогичным образом нарушения свертываемости крови и тромбоз коронарных сосудов имеют свою сложную регуляторную обусловленность. Резкое увеличение функции сердца со склерозированными коронарными сосудами, обусловленное сильными адренергическими эффектами или повышением сопротивления сосудов большого круга кровообращения, составляет не менее частую регуляторную ситуацию, предопределяющую ишемию миокарда. В плане нашего изложения существенно, что многообразные нарушения регуляции, формирующие патогенетическую цепь ишемической болезни и ее исход, нередко протекающий в виде летальных аритмий и острой сердечной смерти, обычно детерминированы чрезмерной по силе и длительности стресс-реакцией организма. В соответствии с этим мы вначале оценим основные патогенетические механизмы, за счет которых стресс-реакция включается в патогенез ишемической болезни, далее специально остановимся на ее роли в патогенезе аритмий, фибрилляции и внезапной сердечной смерти. На этой основе будет представлен главный в данной главе материал о возможности предупреждения аритмий и фибрилляции сердца с помощью адаптации к факторам среды, при которой развивается увеличение мощности и эффективности стресс-лимитирующих систем.
|
|
|
