 |


Транскрипция и трансляция у прокариот
|
|
|
|
Световая репарация
Под влиянием УФ - излучения образуются димеры пиримидиновых оснований. Фермент фотолиаза восстанавливает структуру ДНК (активен под действием света).
Эксцизионная репарация
- это восстановление поврежденного участка ДНК путем его вырезания. Происходит в
1) G1 – периоде (дорепликативная)
2 ) S – периоде (репликативная)
Пострепликативная репарация
- обмен фрагментами (рекомбинация) между двумя вновь образованными спиралями ДНК. Происходит в G2 – периоде клеточного цикла.
SOS – репарация
- достраивание ДНК напротив дефектных звеньев матричной цепи. В результате ДНК оказывается удвоенной, хотя и с большим количеством ошибок, но это дает возможность провести клеточное деление.
Часто становится причиной опасных мутаций, в т.ч. онкологий.
Пример мутации: пигментная ксеродерма – дефект разных репарирующих систем. Возможно снижение активности УФ – эндонуклеазы. До 90% всех раковых заболеваний, возможно, связаны с отсутствием репарации ДНК.
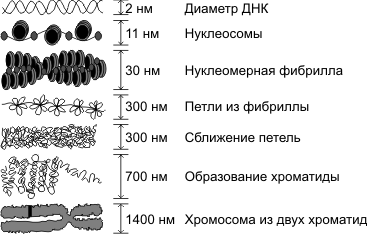
8. Формирование нуклеосомы. Белки, участвующие в формировании её
Нуклеосома — структурная часть хроматина, образованная участком нити ДНК намотанной на сердцевину из основных белков-гистонов, имеет диаметр ~ 11 нм. Нуклеосомы является первым уровнем упаковки ДНК эукариот (требуется для помещения ДНК общей длиной около метра в ядро диаметром 5-10 мкм), а также архей, и обеспечивает ее компактизации примерно в семь раз и защищает ее от повреждений. Сердцевина нуклеосомы состоит из восьми белков: по две копии гистонов H2A, H2B, H3 и H4, N-конец (хвост гистона) каждой белковой молекулы выступает наружу нуклеосомы и может быть местом различных ковалентных модификаций. Длина участка ДНК, входит в состав нуклеосом, составляет 146 п.н., она намотана на белковую сердцевину в виде ливозакрученои суперспирали 1,7 раза. Нуклеосомы размещаются вдоль ДНК достаточно регулярно, на наиболее часто расстоянии от 8 до 80 (длина линкерных ДНК, связанной с гистонов H1). Таким образом нуклеосомы повторяются примерно каждые 200 п.н., одна диплоидная клетка человека содержит примерно 30 млн нуклеосом. Последовательность нуклеосом и линкерных участков формирует 10 нм филамент, имеющий вид «бусин на нитке» и преимущественно подлежит дальнейшей компактизации.
|
|
|
Технически, термин «нуклеосома» обозначает ќорову частичку (146 п.н. ДНК и белковую сердцевину) и одну ближайшую линкерных последовательность, но в основном используется как синоним коревой частицы.
В течение клеточного цикла нуклеосомы временно отсоединяются от ДНК только при репликации, также они могут быть устранены на участках, транскрибируются в определенный момент.
9!!!
Химический состав хромосомы. Классификация хромосом.
Хромосомы
Хромосомы — это цитологические палочковидные структуры, представляющие собой конденсированный хроматин и появляющиеся в клетке во время митоза или мейоза. Хромосомы и хроматин — различные формы пространственной организации дезоксирибонуклеопротеидного комплекса, соответствующие разным фазам жизненного цикла клетки. Химический состав хромосом такой же, как и хроматина: 1) ДНК (30–45%), 2) гистоновые белки (30–50%), 3) негистоновые белки (4–33%).
Основу хромосомы составляет одна непрерывная двухцепочечная молекула ДНК; длина ДНК одной хромосомы может достигать нескольких сантиметров. Понятно, что молекула такой длины не может располагаться в клетке в вытянутом виде, а подвергается укладке, приобретая определенную трехмерную структуру, или конформацию. Можно выделить следующие уровни пространственной укладки ДНК и ДНП: 1) нуклеосомный (накручивание ДНК на белковые глобулы), 2) нуклеомерный, 3) хромомерный, 4) хромонемный, 5) хромосомный.
|
|
|
В процессе преобразования хроматина в хромосомы ДНП образует не только спирали и суперспирали, но еще петли и суперпетли. Поэтому процесс формирования хромосом, который происходит в профазу митоза или профазу 1 мейоза, лучше называть не спирализацией, а конденсацией хромосом.
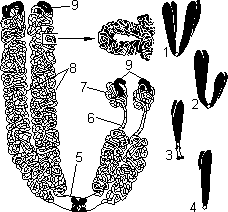
Хромосомы: 1 — метацентрическая; 2 — субметацентрическая; 3, 4 — акроцентрические. Строение хромосомы: 5 — центромера; 6 — вторичная перетяжка; 7 — спутник; 8 — хроматиды; 9 — теломеры.
Метафазная хромосома (хромосомы изучаются в метафазу митоза) состоит из двух хроматид (8). Любая хромосома имеет первичную перетяжку (центромеру) (5), которая делит хромосому на плечи. Некоторые хромосомы имеют вторичную перетяжку (6) и спутник (7). Спутник — участок короткого плеча, отделяемый вторичной перетяжкой. Хромосомы, имеющие спутник, называются спутничными (3). Концы хромосом называются теломерами (9). В зависимости от положения центромеры выделяют: а) метацентрические (равноплечие) (1), б) субметацентрические (умеренно неравноплечие) (2), в) акроцентрические (резко неравноплечие) хромосомы (3, 4).
Соматические клетки содержат диплоидный (двойной — 2n) набор хромосом, половые клетки — гаплоидный (одинарный — n). Диплоидный набор аскариды равен 2, дрозофилы — 8, шимпанзе — 48, речного рака — 196. Хромосомы диплоидного набора разбиваются на пары; хромосомы одной пары имеют одинаковое строение, размеры, набор генов и называются гомологичными.
Кариотип — совокупность сведений о числе, размерах и строении метафазных хромосом. Идиограмма — графическое изображение кариотипа. У представителей разных видов кариотипы разные, одного вида — одинаковые. Аутосомы — хромосомы, одинаковые для мужского и женского кариотипов. Половые хромосомы — хромосомы, по которым мужской кариотип отличается от женского.
Хромосомный набор человека (2n = 46, n = 23) содержит 22 пары аутосом и 1 пару половых хромосом. Аутосомы распределены по группам и пронумерованы:
Половые хромосомы не относятся ни к одной из групп и не имеют номера. Половые хромосомы женщины — ХХ, мужчины — ХУ. Х-хромосома — средняя субметацентрическая, У-хромосома — мелкая акроцентрическая.
|
|
|
В области вторичных перетяжек хромосом групп D и G находятся копии генов, несущих информацию о строении рРНК, поэтому хромосомы групп D и G называются ядрышкообразующими.
Функции хромосом: 1) хранение наследственной информации, 2) передача генетического материала от материнской клетки к дочерним.
11 матричные процессы
Матричными называются такие процессы, при которых на основе первичной структуры одного биополимера, называемой матрицей, синтезируется первичная структура другого биополимера, называемого копией, причем структура матрицы определяет структуру копии. К матричным процессам относятся:
1.биосинтез ДНК или репликация;
2.биосинтез РНК или транскрипция;
3. биосинтез белка или трансляция.
Любой матричный процесс можно разбить на 3 фазы:
1.начало синтеза или инициация
2.продолжение синтеза или элонгация
3.окончание синтеза или терминация.
Это ферментативные процессы, кроме того, требующие затраты не ферментных белковых факторов. Это энергозависимые процессы, которые требуют затраты энергии в виде АТФ или ГТФ. Ведущим правилом всех матричных процессов является правило комплиментарности. В ходе первых двух процессов, которые в основном осуществляются в ядре клетки, матрицей является нуклеиновые кислоты и копией нуклеиновые кислоты. В процессе транскрибции матрицей является нуклеиновая кислота, а копия полипептидная цепь.
Трнскрипция!!!
12 Смысловая и антисмысловая нить ДНК и работа их в транскрипции
Кодирующая цепь (нить), смысловая цепь (нить) (coding chain (strand), sense chain (strand)) [франц. code — код, шифр] — одна из цепей двухцепочечной молекулы ДНК, нуклеотидная последовательность которой идентична мРНК, транскрибированной с комплементарной антисмысловой цепи (матричная цепь ДНК). Иногда обе нити ДНК могут транскрибироваться, но в противоположных направлениях
екодирующая цепь (нить), антисмысловая цепь (нить) (noncoding chain (strand), antisense chain (strand)) [франц. code — код, шифр; греч. anti — против] — одна из двух комплементарных цепей ДНК, которая по последовательности подобна РНК (при условии замены T на У). Общепринято давать структуру гена в виде последовательности Н.ц. ДНК в направлении 5'?3'. Если прочитать кодоны Н.ц. в этом направлении, то с помощью генетического кода можно воспроизвести аминокислотную последовательность белка в принятом порядке, т. е. от N- к С-концу.
|
|
|
11 12 13
Строение гена эукариот
Ген — участок молекулы ДНК, кодирующий первичную последовательность аминокислот в полипептиде или последовательность нуклеотидов в молекулах транспортных и рибосомных РНК. ДНК одной хромосомы может содержать несколько тысяч генов, которые располагаются в линейном порядке. Место гена в определенном участке хромосомы называется локусом. Особенностями строения гена эукариот являются: 1) наличие достаточно большого количества регуляторных блоков, 2) мозаичность (чередование кодирующих участков с некодирующими). Экзоны (Э) — участки гена, несущие информацию о строении полипептида. Интроны (И) — участки гена, не несущие информацию о строении полипептида. Число экзонов и интронов различных генов разное; экзоны чередуются с интронами, общая длина последних может превышать длину экзонов в два и более раз. Перед первым экзоном и после последнего экзона находятся нуклеотидные последовательности, называемые соответственно лидерной (ЛП) и трейлерной последовательностью (ТП). Лидерная и трейлерная последовательности, экзоны и интроны образуют единицу транскрипции. Промотор (П) — участок гена, к которому присоединяется фермент РНК-полимераза, представляет собой особое сочетание нуклеотидов. Перед единицей транскрипции, после нее, иногда в интронах находятся регуляторные элементы (РЭ), к которым относятся энхансеры и сайленсеры. Энхансеры ускоряют транскрипцию, сайленсеры тормозят ее.

Транскрипция у эукариот
Транскрипция — синтез РНК на матрице ДНК. Осуществляется ферментом РНК-полимеразой.
РНК-полимераза может присоединиться только к промотору, который находится на 3'-конце матричной цепи ДНК, и двигаться только от 3'- к 5'-концу этой матричной цепи ДНК. Синтез РНК происходит на одной из двух цепочек ДНК в соответствии с принципами комплементарности и антипараллельности. Строительным материалом и источником энергии для транскрипции являются рибонуклеозидтрифосфаты (АТФ, УТФ, ГТФ, ЦТФ).
В результате транскрипции образуется «незрелая» иРНК (про-иРНК), которая проходит стадию созревания или процессинга. Процессинг включает в себя: 1) КЭПирование 5'-конца, 2) полиаденилирование 3'-конца (присоединение нескольких десятков адениловых нуклеотидов), 3) сплайсинг (вырезание интронов и сшивание экзонов). В зрелой иРНК выделяют КЭП, транслируемую область (сшитые в одно целое экзоны), нетранслируемые области (НТО) и полиадениловый «хвост».
|
|
|

Транслируемая область начинается кодоном-инициатором, заканчивается кодонами-терминаторами. НТО содержат информацию, определяющую поведение РНК в клетке: срок «жизни», активность, локализацию.
Транскрипция и процессинг происходят в клеточном ядре. Зрелая иРНК приобретает определенную пространственную конформацию, окружается белками и в таком виде через ядерные поры транспортируется к рибосомам; иРНК эукариот, как правило, моноцистронны (кодируют только одну полипептидную цепь).
Трансляция
Трансляция — синтез полипептидной цепи на матрице иРНК.
Органоиды, обеспечивающие трансляцию, — рибосомы. У эукариот рибосомы находятся в некоторых органоидах — митохондриях и пластидах (70S-рибосомы), в свободном виде в цитоплазме (80S-рибосомы) и на мембранах эндоплазматической сети (80S-рибосомы). Таким образом, синтез белковых молекул может происходить в цитоплазме, на шероховатой эндоплазматической сети, в митохондриях и пластидах. В цитоплазме синтезируются белки для собственных нужд клетки; белки, синтезируемые на ЭПС, транспортируются по ее каналам в комплекс Гольджи и выводятся из клетки. В рибосоме выделяют малую и большую субъединицы. Малая субъединица рибосомы отвечает за генетические, декодирующие функции; большая — за биохимические, ферментативные.
В малой субъединице рибосомы расположен функциональный центр (ФЦР) с двумя участками — пептидильным (Р-участок) и аминоацильным (А-участок). В ФЦР может находиться шесть нуклеотидов иРНК, три — в пептидильном и три — в аминоацильном участках.
Для транспорта аминокислот к рибосомам используются транспортные РНК, тРНК (лекция №4). Длина тРНК от 75 до 95 нуклеотидных остатков. Они имеют третичную структуру, по форме напоминающую лист клевера. В тРНК различают антикодоновую петлю и акцепторный участок. В антикодоновой петле РНК имеется антикодон, комплементарный кодовому триплету определенной аминокислоты, а акцепторный участок на 3'-конце способен с помощью фермента аминоацил-тРНК-синтетазы присоединять именно эту аминокислоту (с затратой АТФ). Таким образом, у каждой аминокислоты есть свои тРНК и свои ферменты, присоединяющие аминокислоту к тРНК.

| 
| |
| 1 — антикодон; 2 — участок, связывающий аминокислоту. | Транспорт аминокислот к рибосомам: 1 — фермент; 2 — тРНК; 3 — аминокислота. |
Двадцать видов аминокислот кодируются 61 кодоном, теоретически может быть 61 вид тРНК с соответствующими антикодонами. Но кодируемых аминокислот всего 20 видов, значит, у одной аминокислоты может быть несколько тРНК. Установлено существование нескольких тРНК, способных связываться с одним и тем же кодоном (последний нуклеотид в антикодоне тРНК не всегда важен), поэтому в клетке обнаружено всего около 40 различных тРНК.

Синтез белка начинается с того момента, когда к 5'-концу иРНК присоединяется малая субъединица рибосомы, в Р-участок которой заходит метиониновая тРНК (транспортирующая аминокислоту метионин). Следует отметить, что любая полипептидная цепь на N-конце сначала имеет метионин, который в дальнейшем чаще всего отщепляется. Синтез полипептида идет от N-конца к С-концу, то есть пептидная связь образуется между карбоксильной группой первой и аминогруппой второй аминокислот.
Затем происходит присоединение большой субъединицы рибосомы, и в А-участок поступает вторая тРНК, чей антикодон комплементарно спаривается с кодоном иРНК, находящимся в А-участке.
Пептидилтрансферазный центр большой субъединицы катализирует образование пептидной связи между метионином и второй аминокислотой. Отдельного фермента, катализирующего образование пептидных связей, не существует. Энергия для образования пептидной связи поставляется за счет гидролиза ГТФ.
Как только образовалась пептидная связь, метиониновая тРНК отсоединяется от метионина, а рибосома передвигается на следующий кодовый триплет иРНК, который оказывается в А-участке рибосомы, а метиониновая тРНК выталкивается в цитоплазму. На один цикл расходуется 2 молекулы ГТФ. В А-участок заходит третья тРНК, и образуется пептидная связь между второй и третьей аминокислотами.
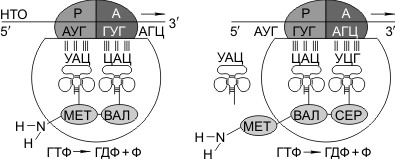
Трансляция идет до тех пор, пока в А-участок не попадает кодон-терминатор (УАА, УАГ или УГА), с которым связывается особый белковый фактор освобождения. Полипептидная цепь отделяется от тРНК и покидает рибосому. Происходит диссоциация, разъединение субъединиц рибосомы.
Скорость передвижения рибосомы по иРНК — 5–6 триплетов в секунду, на синтез белковой молекулы, состоящей из сотен аминокислотных остатков, клетке требуется несколько минут. Первым белком, синтезированным искусственно, был инсулин, состоящий из 51 аминокислотного остатка. Потребовалось провести 5000 операций, в работе в течение трех лет принимали участие 10 человек.
В трансляции можно выделить три стадии: а) инициации (образование иницаторного комплекса), б) элонгации (непосредственно «конвейер», соединение аминокислот друг с другом), в) терминации (образование терминирующего комплекса).
Транскрипция и трансляция у прокариот
«Механизмы» сборки полинуклеотидных и полипептидных цепочек у прокариот и эукариот не различаются. Но в связи с тем, что гены прокариот не имеют экзонов и интронов (исключение — гены архебактерий), располагаются группами, и на эту группу генов приходится один промотор, появляются следующие особенности транскрипции и трансляции у прокариот.
- В результате транскрипции образуется полицистронная иРНК, кодирующая несколько белков, совместно обеспечивающих определенную группу реакций.
- иРНК имеет несколько центров инициации трансляции, терминации трансляции и НТО.
- Не происходят КЭПирование, полиаденилирование и сплайсинг иРНК.
- Трансляция начинается еще до завершения транскрипции; эти процессы не разделены во времени и пространстве, как это имеет место у эукариот.
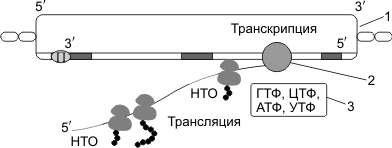
1 — ДНК; 2 — РНК-полимераза; 3 — Нуклеозидтрифосфаты ГТФ, ЦТФ, АТФ, УТФ.
Можно добавить, что срок «жизни» прокариотических иРНК — несколько минут (у эукариот — часы и даже сутки).
14. Генетический код. Свойства генетического кода и их характеристика.
|
|
|
