 |


Чувствительнык нервные окончания
|
|
|
|
Лекция №
Нервные окончания
Все нервные волокна заканчиваются нервными окончаниями. По функции нервные окончания делят на три группы:
1. межнейронные синапсы ( контакты) – обеспечивают функциональную связь между нейронами;
2. эффекторные ( эфферентные) окончания – передают сигналы из нервной системы на исполнительные органы (мышцы, железы), имеются на аксонах;
3. аффекторные ( рецепторные или чувствительные) окончания – воспринимают раздражения из внешней и внутренней среды, имеются на дендритах.
Межнейронные синапсы – это специфические контакты между нервными клетками. Межнейральные синапсы подразделяют на электрические и химические.
Электрические синапсы – имеют вспомогательное значение. Пологают, что в эволюции они послужили той основой, на которой развивались химические синапсы. В электрических синапсах импульс может передаваться в обоих направлениях. Но есть специализированные синапсы, в которых передача может осуществляться в одном направлении.
Электрические синапсы существуют в двух разновидностях: 1-с синаптической щелью и 2-без нее. В первом случае мембраны синаптически связанных нейронов разделены промежутком шириной 2 нм, пронизанным коннексонами.. Коннексоны представляю собой трубочки, образованные молекулами интегральных белков, выступающих над липидным слоем мембраны клеток и служат водными каналами, через которые мелкие молекулы и ионы транспортируются из одной клетки в другую. Когда потенциал действия, распространяющийся по мембране одной клетки достигает области щелевого соединения, электрический ток пассивно протекает через щель от одной клетки к другой.
Во второй разновидности щель между мембранами отсутствует и они непосредственно соприкасаются своими нейролеммами.
|
|
|
Химический тип синапсов – наиболее характерен и специфичен для нервной системы млекопитающих. Действие химических синапсов основано на преобразовании электрического сигнала в химический, который затем вновь преобразуется в электрический.
По локализации различают: аксо-соматические – терминали одного нейрона оканчиваются на теле другого, аксо-дендрические терминали аксона одного нейрона вступают в контакт с дендритом другого, и аксо-аксональные – терминали аксона одного нейрона оканчиваются на аксонах другого.
Химические синапсы обеспечивают одностороннюю передачу сигнала. Взаимоотношения между клетками при работе химических синапсов происходит по принципу «команда-исполнитель», а не по принципу товарищеского взаимодействия как в случае электрических синапсов. Химический синапс состоит из 3-х компонентов: пресинаптической части ( принадлежит командующей клетке), постсинаптической части (принадлежит клетке исполнительнице) и синаптической щели.
Пресинаптическая часть образуется аксоном, в ней содержатся митохондрии, аЭПС, нейрофиламенты, нейротрубочки и синаптические пузырьки (диаметром от 20-65 нм), содержащие нейромедиатор. В качества медиатора в разных синапсах используются одно из соединений: ацетилхолин, холин, норадреналин, серотонин, различные полипептиды, аминокислоты и т.д. Форма, размер синаптических пузырьков и электронная плотность их содержимого зависит от степени их зрелости и химической природы медиатора. В холинэргических синапсах они мелкие и прозрачные, в адренергических – крупные с плотной гранулой. Нейромедиаторы синтезируются в теле нейрона и механизмом быстрого транспорта переносятся в окончание аксона, где депонируются. Частично синаптические пузырьки образуются в самом синапсе путем отщепления от цистерн аЭПС. Под влиянием нерного импульса нейромедиатор выделяется в синаптическую щель через синаптические утолщения, т.н. активные зоны, имеющиеся на пресинаптической мембране.
|
|
|
Постсинаптическая часть представленапостсинаптической мембраной, которая содержит синаптические рецепторы (особые комплексы интегральных мембран), которые связываются с медиаторами.
Синаптическая щель шириной 20-30 нм иногда содержит гликопротеиновые филаменты, которые являются элементами специализированного гликокаликса и обеспечивают адгезивные связи между пре- и постсинаптической частями и направленную диффузию медиатора.
Механизм передачи нервного импульса в химическом синапсе.
Под влиянием нервного импульса потенциалзависимые кальциевые каналы пресинаптической мембраны активируются, что вызывает приток ионов Са в аксон. В присутствии ионов Са мембраны синаптических пузырьков сливаются с пресинаптической мембраной и их содержимое (медиатор) выделяется в синаптическую щель механизмом экзоцитоза. Медиатор связывается с рецепторами в постсинаптической части, что приводит к ее деполяризации и возникновению нервного импульса в возбуждающих синапсах, либо гиперполяризации, вызывая реакцию торможения в тормозных синапсах.
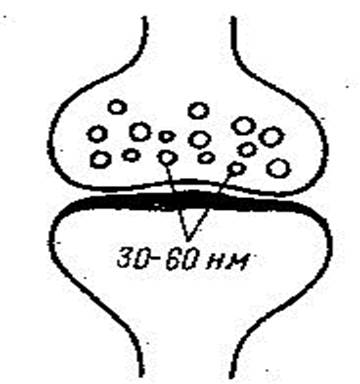
В отсутствии нервного импульса пресинаптическая часть выделяет небольшие порции медиатора, которые вызывают в постсинаптической мембране спонтанные небольшие потенциалы.
Эффекторные нервные окончания – это концевые аппараты аксонов двигательных нейронов. Они двух типов двигательные и секреторные. Двигательные окончания имеются в поперечнополосатых и гладких мышцах, секреторные – в железах.
Двигательное окончание на поперечнополосатых соматических мышцах называется нервно-мышечным синапсом или моторной бляшкой. Это окончания аксонов нейронов двигательных ядер передних рогов спинного мозга и моторных ядер головного мозга. В двигательном окончании выделяют три части: пресинаптическую часть – это концевые ветвления аксона, постсинаптическую часть – специализированный участок на мышечном волокне и синаптическую щель – располагается между ветвлениями аксона и мышечным волокном.
|
|
|
В крупных мышцах, развивающих большую силу, один аксон разветвляясь, может иннервировать сотни и тысячи мышечных волокон. В мелких мышцах, осуществляющих тонкие движения (например, наружных мышц глаза), каждое волокно или небольшая их группа иннервируются отдельным аксоном.
Пресинаптическая часть. Миелиновое волокно подойдя к мышечному теряет миелиновую оболочку и дает несколько терминалей, которые погружаются в мышечное волокно, увлекая за собой его плазмолемму. Веточки аксона с поверхности покрыты уплощенными леммоцитами и базальной мембраной, которая переходит с мышечного волокна. В терминалях аксона находятся митохондрии и синаптические пузырьки, которые содержат ацетилхолин (медиатор). Аксолемма нервной терминали в области синапса является пресинаптической мембраной. Она содержит потенциалзависимые Са каналы. При деполяризации мембраны каналы открываются и ионы Са входят в терминаль, запуская секрецию квантов ацетилхолина.
Синаптическая щель. Ширина щели 50 нм. Она содержит базальную мембрану и отростки глиальных клеток. Базальная мембрана удерживает в области синапса терминали аксона, контралирует расположение рецепторов в постсинаптической мембране. Она содержит несколько сигнальных белков (агрин, S-ламинин и др), которые служат метками при помощи которых регенирирующий аксон находит синаптическую область на поперечнополосатом мышечном волокне.
Постсинаптическая часть. Плазмолемма мышечного волокна в области синапса является постсинаптической мембраной. Она образует многочисленные инвагинации и вторичные складки за счет которых сильно увеличивается площадь мембраны. В мембрану встроены холинорецепторы (20-30 тыс. на 1 кв.мкм). Мышечное волокно в области синапса не имеет исчерченности, содержит многочисленные митохондрии, цистерны грЭПС, рибосомы и скопления ядер.
Механизм передачи нервного импульса на мышечное волокно в нервно-мышечном синапсе сходен в химическом межнейральном синапсе.
|
|
|
Синаптическая передача включает ряд последовательных процессов:
1 – синтез нейромедиатора. Происходит это в терминали, ферменты необходимые для его синтеза образуются в перикарионе и поступают в терминали.;
2 – хранение нейромедиатора. Накапливается и хранится в нервной терминали, находясь в синаптических пузырьках вместе с АТФ и некоторыми катионами. В пузырьке находится несколько тысяч молекул нейромедиатора, что составляет квант.
3 – секреция нейромедиатора. При достижении потенциала действия нервной терминали концентрация ионов Са в цитозоли резко увеличивается, что приводит к экзоцитозу квантов нейромедиатора в синаптическую щель.
4 – взаимодействие медиатора с рецепторами, встроенными в постсинаптичесую мембрану, что вызывает ее деполяризацию и последующее сокращение мышечного волокна.
5 – удаление нейромедиатора. Медиатор отщепляется от рецептора и быстро разрушается ферментом ацетилхолинэстеразой, который содержится в синаптической щели.
Чувствительнык нервные окончания
Тельца Руффини – крупные веретеновидной формы структуры длиной 2 мм, диаметром 150 мкм, расположены в соединительной ткани кожи и суставов. Тельце содержит внутреннюю колбу с густой сетью разветвленных нервных терминалей с митохондриями, везикулами и вспомогательными пластинчатыми клетками. Снаружи тельце покрыто соединительнотканной капсулой из нескольких слоев уплощенных фибробластов. Между внутренней колбой и капсулой находится заполненное жидкостью капсулярное пространство.
Колба Краузе – мелкие округлые тельца, являются механорецепторами и предположительно холодовыми рецептора. Содержатся в соединительной ткани сосочкового слоя кожи и собственной пластинке слизистой оболочки ротовой полости, надгортаннике, в коньюктиве глаза. Внутренняя колба образована уплощенными глиальными клетками, между которыми тонкие терминали дендрита образуют сплетение в виде клубочка. Капсула состоит из плоских клеток.
Пластинчатые (или фатер-пачиниевы) тельца. Находятся в глубоких слоях дермы и в строме внутренних органов. Воспринимают относительно сильное давление. Состоит из тех же трех элементов: дендрита, внутренней колбы и соединительнотканной оболочки.
1. Терминали дендрита (лишенные миелиновой оболочки) - располагаются внутри тельца. 2. Окружающие их глиальные клетки - образуют т.н. внутреннюю колбу (или внутреннюю луковицу).
3. Соединительнотканная оболочка, или наружная колба (наружная луковица) образована плотной волокнистой соединительной тканью, состоящей из фибробластов и спирально ориентированными коллагеновыми волокнами. Она толстая, имеет пластинчатую структуру, т.е. содержит несколько слоёв. Вследствие такого её строения, пластинчатое тельце воспринимает достаточно сильные давления.
|
|
|
В мышцах и сухожилиях рецепторные окончания тоже являются инкапсулированными.
Они представляют собой особые структуры - нервно-мышечные и
нервно-сухожильные веретёна. Нервно-мышечные веретёна располагаются в толще скелетных мышц. Веретено (fusus) содержит 4 компонента: от 1 до 12 специальных (т.н. интрафузальных ) мышечных волокон (1), растяжимую соединительнотканную капсулу (2) вокруг веретена, афферентные нервные волокна (3) и их окончания, которые под капсулой оплетают центральные части интрафузальных волокон.
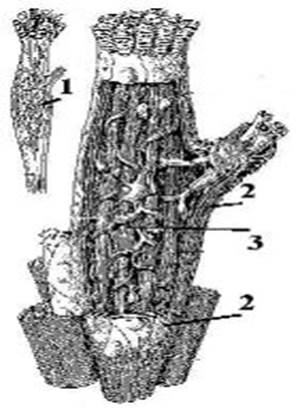
На концах этих волокон оканчиваются эфферентные нервные волокна, идущие от гамма-мотонейронов спинного мозга. У интрафузальных волокон сокращаются только концы, содержащие миофибриллы, а центральные части их лишены и они являются рецепторными. Интрафузальные волокна двух типов: 1- с ядерной сумкой, в них ядра образуют плотное скопление в экватариальной области; 2 - с ядерной цепочкой, они тоньше и короче первых и ядра в них располагаются в виде цепочки.
К интрафузальным мышечным волокнам подходят нервные аферентные волокна двух типов. Первичные волокна (17 мкм) образуют спираль в пределах экваториальной зоны обоих типов интрафузальных мышечных волокон. Вторичные волокна (8 мкм) иннервируют волокна с ядерной цепочкой рядом с экваториальной зоной. Волокна с ядерной сумкой реагируют на скорость растяжения,а волокна с ядерной цепочкой - на общую величину растяжения.
Нервно-сухожильные веретена расположены в концевой части сухожилия на границе с мышцей и в связках капсулы суставов. Рецептор имеет веретеновидную форму и окружен соединительнотканной капсулой.
Миелиновое волокно подходит к нервно-сухожильному веретену теряет миелиновую оболочку и образует терминали, которые ветвятся между пучками коллагеновых волокон сухожилия. Они предотвращают перерастяжение мышц.
|
|
|
