 |


Развитие и размножение половых клеток
|
|
|
|
ВВЕДЕНИЕ
В учебном плане подготовки бакалавров и специалистов биологов важное место отводится биологии размножения и развития (БРР). Целью данной дисциплины является выяснение закономерностей предзародышевого развития (т.е. формирования половых клеток), всех процессов зародышевого развития, причин аномалий и возникновения уродств. Важное научно-практическое значение БРР связано с получением новых знаний о размножении и развитии на молекулярном уровне и их применении для решения актуальных проблем медицины и животноводства, остро стоящих перед специалистами в связи с развитием технологий экстракорпорального оплодотворения у людей и искусственного осеменения у сельскохозяйственных животных.
В связи с этим подчеркнем, что изучение половых клеток млекопитающих является важной проблемой современной биологии. Полученные данные позволяют дополнять и расширять теоретическую базу медицины и животноводства, развивать имеющие и создавать новые инструменты для решения практических задач, связанных с репродукцией. Знание частных вопросов гаметогенеза позволяет селекционерам выбирать эффективный генетический материал для создания новых пород животных. Ранняя диагностика морфофункциональных изменений половых клеток позволяет избежать нежелательных последствий, дает возможность предотвратить рождение аномального потомства.
В настоящее время все большую социальную и медицинскую значимость приобретает проблема бесплодного брака. Нарушение оплодотворяющей способности мужчин оказывается причиной бесплодного брака в 40 − 50% наблюдений. Одним из важнейших методов в оценке функционального состояния половых желез и плодовитости мужского пола является исследование спермы. Специальные методы исследования половых клеток играют важную роль в постановке и формировании окончательного диагноза.
|
|
|
В животноводстве все большее распространение получает искусственное осеменение животных, эффективность которого зависит от значительного числа факторов, зависимых от активности сперматозоидов.
В настоящих рекомендациях рассматриваются теоретические и практические аспекты изучения половых клеток, позволяющие ознакомиться с проблемами, возникающими перед биологами, медиками и зоотехниками, изучающими и использующими сперматозоиды в своей работе, а также найти некоторые пути их решения.
ТЕОРЕТИЧЕСКАЯ ЧАСТЬ
Развитие и размножение половых клеток
Первичные половые клетки. Клетки организма подразделяются на половые и соматические. В отличие от соматических клеток, половые гаметы (от греч. gametes – супруг) специализированы на воспроизводстве поколений организмов и имеют половинный (гаплоидный) генетический набор (lcln, или 23 хромосомы – у человека). Различают мужские и женские половые клетки, которые несут генетическую информацию по отцовской и материнской линиям. В половых клетках у человека присутствуют 22 аутосомы и 1 половая хромосома, которая обозначается как X или Y – у мужчин и X – у женщин. При слиянии мужской и женской половых клеток в процессе оплодотворения образуется диплоидная клетка – зигота, дающая начало всем клеткам нового организма. При первом знакомстве со строением зрелых мужских и женских половыхклеток обращает на себя внимание их высокая функциональная целесообразность. У большинства организмов (насекомые, позвоночные и другие) обособление половых клеток происходит на ранних этапах эмбриогенеза. В отличие от соматических клеток, половые клетки тотипотентны, т.е. сохраняют способность формировать все органы и ткани организма. Половые клетки у животных, имеющих морфологически выраженные гонады, закладываются вне гонады. С момента выделения и до вселения в гонаду их называют первичными половыми клетками (ППК). У большинства животных ППК – единственный источник зрелых гамет. ППК попадают в область зачатка гонады с током жидкости или активно мигрируя через клеточные пласты на большие расстояния посредством амебовидного движения. Миграция клеток может происходить посредством гаптотаксиса, контактного ориентирования и хемотаксиса. У некоторых видов ППК можно распознать по своеобразной морфологии (размеры, форма) либо по специфическим гистохимическим реакциям окрашивания цитоплазмы. Так, для ППК млекопитающих и птиц характерен высокий уровень активности щелочной фосфатазы. Используя метод выявления этого фермента можно проследить путь перемещения ППК вплоть до момента проникновения в гонады. Первоначально ППК присутствуют в зародыше в небольшом количестве, но они проходят ряд делений и число их возрастает. У зародышей мыши первые ППК выявляются в стенке аллантоиса и в прилегающем желточном мешке. Считается, что в этот момент ППК уже разделились на две группы, которые будут мигрировать в правую и левую гонады. Затем ППК двигаются вверх по дорсальной брыжейке, на 10 – 11-е сутки они достигают закладки гонады и дают начало линии стволовых половых клеток в гонаде. ППК могут нормально дифференцироваться в половые клетки только в том случае, если попадут в половые валики и вступят в контакт с соматическими клетками гонад, получая при этом необходимую информацию для последующей дифференцировки в оогонии или сперматогонии. Если же на своем пути ППК оказываются в окружении других зачатков и не достигают гонад, то либо гибнут, либо становятся источником тератом. Мужские и женские ППК неотличимы. Несмотря на то, что пол зародыша определяется хромосомным набором, полученным зиготой при оплодотворении, различия в их строении становятся заметными лишь при дифференцировке половой железы. Зачатки мужских и женских гонад состоят из корковой и мозговой области. В случаях развития семенников или яичников возникают различия в положении половых клеток; у самцов они располагаются в мозговой, а у самок – в корковой области, давая начало соответственно сперматогониям либо оогониям.
|
|
|
Гаметогенез. Развитие половых клеток – гаметогенез – происходит в семенниках или яичниках. Процесс развития мужских половых клеток называется сперматогенезом, женских половых клеток – оогенезом.
|
|
|
Сперматогене́з (от греч. sperma, spermatos – семя, genesis – происхождение, возникновение) представляет собой превращение первичных мужских половых клеток в зрелые сперматозоиды и происходит регулирующим влиянием гормонов. Сперматогенез можно разделить на три основные стадии: размножение клеток путем митоза, мейоз и спермиогенез. Митоз первичных половых клеток происходит на протяжении всей жизни особи мужского пола, начиная с периода полового созревания. Митотически активные клетки, находящиеся вблизи наружной стенки семенных канальцев, известны под названием сперматогоний. Сперматогонии А представляют собой популяцию стволовых клеток. В ней имеется группа темных неделящихся клеток, которые, вероятно, образуют «долгосрочный резерв». Некоторые из этих клеток превращаются в митотически активные бледные клетки, а в конечном счете – в сперматогонии В, которые делятся митотически, дифференцируются в сперматоциты 1-го порядка и вступают в мейоз. Сперматогонии В не всегда располагаются на базальной мембране. Они отличаются от сперматогоний своими округлыми ядрами, содержащими конденсированный хроматин, располагающийся либо вблизи нуклеолеммы, либо вблизи ядрышка. В их цитоплазме содержатся свободные рибосомы, митохондрии, достаточно хорошо выраженный комплекс Гольджи, гранулярный эндоплазматический ретикулум. Для превращения светлой сперматогонии в сперматогонию требуются четыре последовательных митотических деления, при которых генетически клетки не меняются, но происходит их морфофункциональная перестройка. После того как в этих клетках пройдет последнее самоудвоение ДНК, они получают название прелептотенных сперматоцитов 1-го порядка, имеющих большое ядро с хорошо видимыми обособленными хромосомами, по-прежнему содержащее их полный диплоидный набор; теперь они готовы к прохождению мейотической стадии сперматогенеза. Сперматоциты 1-го порядка, или первичные сперматоциты, соединены между собой межклеточными мостиками, которые остаются при митозе как сперматогонии А благодаря неполному разделению клеток. Последующие поколения клеток также остаются соединенными между собой, в результате чего образуется синцитий, клетки которого составляют клон. Клетки синцития делятся синхронно, лишь единичные клетки не делятся. Во время первого деления мейоза (мейоз I) каждый такой сперматоцит 1-го порядка делится на две дочерние клетки. В это время на гистологическом препарате можно наблюдать характерные фигуры разделяющихся хромосом на стадиях метафазной пластинки, анафазы, обособления ядер и образования перетяжки клеток. В результате мейоза I образуются две дочерние клетки – сперматоциты 2-го порядка, каждый из которых содержит гаплоидный набор d-хромосом (d-хромосома – хромосома после репликации ДНК, которая состоит из двух дочерних молекул ДНК). Вторичные сперматоциты расположены ближе к просвету канальца. Во время второго деления мейоза (мейоз II) из каждого сперматоцита 2-го порядка образуются две сперматиды. Таким образом, в результате деления одной сперматогонии образуются четыре сперматиды, каждая из которых обладает гаплоидным набором хромосом (рис. 1).
|
|
|
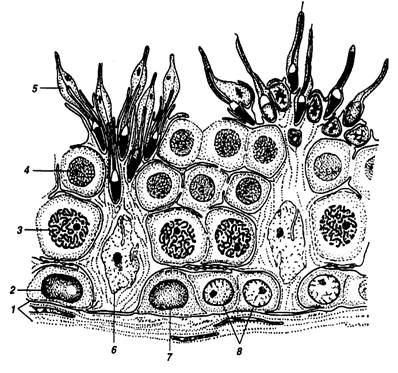
Рисунок 1 – Участок эпителия семенных канальцев, на котором показана связь между клетками Сертоли и развивающимися сперматозоидами:
1 – ограничивающая мембрана; 2 – темная сперматогония А; 3 – сперматоциты в середине пахитены; 4 – сперматиды на ранних стадиях развития; 5 – сперматиды в конце развития; 6 – клетки Сертоли; 7 – бледная сперматогония А; 8 – сперматогонии В
(Б.Карлсон; 1983)
Сперматида – мелкая клетка округлой формы с крупным ядром, которое впоследствии приобретает овоидную форму и располагается эксцентрично (ближе к стенке канальца). Процесс первого мейотического деления занимает несколько недель, тогда как второе завершается в течение 8 ч, поэтому сперматоциты 2-го порядка найти на препаратах трудно. В результате мейотической фазы сперматогенеза образуются четыре гаплоидные сперматиды, содержащие по 22 соматические и 1 половой хромосоме у человека. В отличие от образования женских половых клеток, где всегда образуются гаметы, содержащие женскую Х-хромосому, при сперматогенезе образуются различные сочетания аутосомных и половых хромосом: половина клеток содержит половую Х-хромосому, половина – Y-хромосому.
|
|
|
Таким образом, сперматозоиды можно условно разделить на «женские» (X) и «мужские» (Y): первые после оплодотворения дадут женский пол (XX),
вторые – мужской (XY). Суммарно из одной стволовой сперматогонии образуются 64 сперматиды. По мере созревания клетки продвигаются от базальной мембраны к просвету канальца, и в различных участках канальца можно наблюдать сперматогенные клетки на разных этапах развития и дифференцировки (рис. 1, 2).
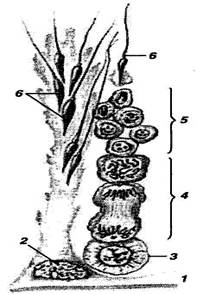
Рисунок 2 – Схема сперматогенеза:
1 – стенка извитого семенного канальца; 2 – поддерживающий эпителиоцит, или сустенциоцит (клетка Сертоли); 3 – сперматогонии; 4 – сперматоциты; 5 – сперматиды; 6 – сперматозоиды (по И. В. Алмазову и Л. С. Сутулову)
Важная особенность сперматогенеза заключается в том, что в ходе последовательных делений цитокинез (расхождение клеток) не доходит до конца. Клетки остаются связанными между собой посредством цитоплазматических мостиков диаметром 1 − 2 мкм, образуя синцитий. Поскольку молекулы и ионы легко проникают через межклеточные мостики, клетки такого клона развиваются синхронно. Такая связь нарушается только при достижении ими стадии сперматид.
Генетически сперматиды представляют собой конечный продукт мейоза. Но, прекращая деление, они подвергаются глубоким преобразованиям, превращаясь из относительно обычных на вид клеток в чрезвычайно специализированные – сперматозоиды.
Третий период сперматогенеза – превращение сперматид в сперматозоиды – носит название спермиогенез, или метаморфоз сперматид. В ходе сложного процесса спермиогенеза сперматиды дифференцируются в зрелые сперматозоиды. Дифференцирующие сперматиды лежат в углублениях плазматической мембраны клеток Сертоли. При спермиогенезе область ядра, обращенная к стенке семенного канальца, постепенно вытягивается и заостряется. Комплекс Гольджи формирует гранулы, которые сливаются между собой, образуя акросомальную гранулу (акробласт), контактирующую с апикальной частью ядра. Одновременно гиперплазирующиеся мешочки комплекса Гольджи окружают акросомальную гранулу, в результате чего образуется акросома, содержащая протеолитические ферменты (в основном гиалуронидазу и трипсиноподобный акрозин), которые при контакте с яйцеклеткой разрушают ее прозрачную зону. При отсутствии или недоразвитии акросомы сперматозоид теряет способность оплодотворять яйцеклетку.
Из одной центриоли начинают образовываться микротрубочки, группирующиеся сначала по три, а затем по две, к ним мигрируют митохондрии, образующие вокруг микротрубочек характерную спиральную структуру. Формируется специализированная органелла движения – жгутик. Одновременно большинство внутриклеточных структур редуцируется. В результате примерно за 16 дней круглая сперматида со сферическим ядром превращается в чрезвычайно специализированную клетку, имеющую маленькую головку длиной 5 мкм и шириной 3 мкм, подвижный хвост длиной 50 мкм, предназначенный для единственной задачи – доставки наследственного материала отца в материнскую яйцеклетку.
В процессе спермиогенеза существенно уменьшается количество цитоплазмы формирующегося сперматозоида. Большая часть ее образует остаточное тельце, которое отделяется от клетки и фагоцитируется поддерживающим эпителиоцитом, или сустентоцитом (клеткой Сертоли). Меньшая часть цитоплазмы покрывает тонким слоем ядро, связующую, промежуточную и не полностью главную части сперматозоида. Разделение клеток одного клона происходит на последних этапах формирования сперматозоидов, которые отделяются от клеток Сертоли и попадают в просвет канальца, где находится жидкость, вырабатываемая клетками Сертоли.
Сперматозоид – мужская половая клетка. Учеловека длиной около
65 мкм, имеет головку и жгутик (хвостовой конец), покрыт, словно чехлом, цитоплазматической мембраной, удлиненная и закругленная (спереди) головка (длина около 5 мкм, ширина около 3 мкм и толщина около 1 мкм) заполнена в основном ядром. С точки зрения гидродинамики, головка наиболее приспособлена для выполнения функции продвижения в жидкости половых путей женщины, проникновения в яйцеклетку и внесения в нее ДНК. Плотное ядро заполнено конденсированным хроматином, лишь некоторые участки его имеют более светлый вид – это ядерные пузырьки. Почти все ядро сперматозоида окутано акросомой, отделенной узкой щелью от цитолеммы и наружной ядерной мембраны. Ядерные поры имеются лишь в дистальном отделе ядра в области связующей части. Уплощенная, несколько вогнутая дистальная часть ядра, лежащая на базальной чашечке – тонкой пластинке мелкозернистого электронно-плотного вещества, образует сочленяющую ямку. На ультрамикроскопическом уровне в головке сперматозоида различимы акросомальная и постакросомальная зоны, в хвосте – промежуточный, главный и концевой отделы. Большую часть головки занимает ядро, заполненное конденсированным хроматином. Пространство между ядром и передним участком плазмолеммы спермия занято акросомальной вакуолью – специализированной лизосомой, содержащей группу ферментов – лизинов оболочек яйцеклетки: акрозин (разрушает прозрачную зону овоцита), пенетразу (диссоциирует клетки лучистого венца), гиалуронидазу (расщепляет гиалуроновую кислоту), кислую фосфатазу (разрушает форсхолин при прохождении спермиев через плазмолемму овоцита). Шейка спермия представляет собой короткий отдел, в котором находятся проксимальная центриоль и 9 сегментированных колонн. Жгутик, или хвостовой конец, сперматозоида состоит из связующей, промежуточной, главной и конечной (терминальной) части. Связующая часть длиной около 1 мкм, начинается базальной чашечкой, содержит проксимальную и дистальную центриоли, окруженные столбчатым основанием, образованным девятью сегментированными полосатыми столбами, являющимися отростками плотных столбов. Проксимально полосатые столбы формируют головку, которая контактирует с базальной чашечкой. От промежуточной части через весь жгутик проходит аксонема, состоящая из девяти периферических двойных и одной пары центральных микротрубочек. Каждая периферическая двойная трубочка состоит из более электронно-плотной трубочки А, снабженной двумя «ручками», и менее плотной трубочки В, лишенной «ручек». От каждой трубочки А к центральной паре отходит полоска осмиофильного вещества, содержащего сократительный белок динеин.
Через промежуточную часть длиной около 7 мкм и толщиной около
1 мкм вокруг аксонемы проходят девять плотных продольно ориентированных волокон, окруженных митохондриальным влагалищем, образованным спирально расположенными митохондриями с хорошо развитыми кристами.
Митохондрии синтезируют АТФ, необходимую для движения жгутика. До сих пор окончательно не выяснена роль плотных волокон. Возможно, они вместе с динеином участвуют в осуществлении движений сперматозоида. Плотное кольцо, связанное с цитолеммой, находится на границе между промежуточной и главной частями сперматозоида. На этом уровне заканчивается митохондриальное и начинается волокнистое влагалище. Промежуточный отдел содержит аксонему и 9 продольно ориентированных элементов цитоскелета, состоящих из кератиноподобных белков и являющихся продолжением сегментированных колонн. Кнаружи от этих волокон располагаются митохондрии. Аксонема построена по принципу реснички, биения которой инициируются катионами кальция, находящимися в окружающей среде, и митохондриальной АТФ спермия. Через главную часть жгутика длиной около 45 мкм и толщиной около 0,5 мкм проходит аксонема, окруженная лежащим под цитолеммой волокнистым влагалищем. Последнее представляет собой тонкофибриллярный чехол, состоящий из двух продольных столбов, расположенных друг против друга, от которых в обе стороны отходят циркулярно расположенные фибриллы. В проксимальной половине главной части между волокнистым влагалищем и аксонемой проходит семь плотных волокон, остальные два трансформировались в продольные столбы волокнистого влагалища. При переходе проксимальной половины главной части в дистальную плотные волокна заканчиваются, и далее аксонема окружена лишь волокнистым влагалищем, которое простирается до начала конечной (терминальной) части, образованной покрытой цитолеммой аксонемой. В самом конце жгутика описанная структура аксонемы нарушается и количество микротрубочек уменьшается. В концевом отделе хвоста элементы цитоскелета исчезают, и аксонема прикрыта лишь плазмолеммой. У человека дуплеты аксонемы в концевом отделе распадаются на 18 одиночных микротрубочек (рис. 3).
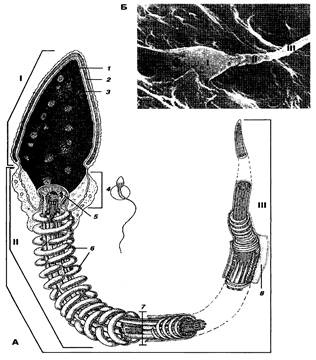
Рисунок 3 – Строение сперматозоида. А – схема, Б – электронная микрофотография; I – головка; II – средняя часть; III – жгутик; 1 – ядро, 2 – акросома; 3 – ядерная оболочка; 4 – шейка; 5 – полосатые столбы; 6 – митохондрии; 7 – аксонема; 8 – цитоплазма
Суммарно у человека развитие сперматозоидов происходит в течение 74 (73 − 75) суток, и каждый час их в яичках образуется около 100 млн штук. Однако сперматозоид, образовавшийся в яичке, еще не способен оплодотворять: жгутик пока не может самостоятельно двигаться и ферменты акросомы не «созрели» для проникновения через защитные оболочки яйцеклетки. Эти свойства сперматозоид приобретает в результате перехода через придаток яичка. Общая длина спермиев составляет 50 − 70 мкм, средний объем 16 − 19 мкм. Для каждого вида животных время, необходимое для превращения сперматогония в зрелый спермий (включая время пребывания в придатке) постоянно, хотя различия между видами существенны. Продолжительность сперматогенеза составляет: у быка – 54 дня; у хряка – 34 дня; у жеребца – 42. Сперматозоиды, закончившие формирование, попадают в систему семявыводящих путей. При совокуплении освобождаются спермии не прямо из семенника, а из каудальной части ("хвоста") придатка семенника. В канале придатка спермии накапливаются в больших количествах (20 − 40 млрд у быка). Здесь они претерпевают дальнейшие морфофункциональные изменения ("дозревают") в течение 8 − 20 дней. В кислой безкислородной среде канала придатка спермии впадают в состояние, подобное анабиозу, приобретают уплотненную липопротеидную оболочку и отрицательный заряд, что предохраняет их от действия кислых продуктов и от агглютинации в половых путях самки. Оплодотворяющую способность спермии сохраняют в придатках семенника до 2 − 3 месяцев. Достигшие каудального отдела придатка спермии обладают высокой оплодотворяющей способностью и могут высвобождаться при эякуляции. По выходе из яичка спермии морфологически сформированы, но функционально незрелы – они неподвижны, не способны к оплодотворению и, помимо этого, являются носителями ряда специфических антигенов, инактивация которых (как и приобретение подвижности и оплодотворяющей способности – капацитации) происходит в системе семявыносящих путей мужского тракта.
Оогенезом называется дифференцировка половых клеток по женскому типу. Развитие женских половых клеток у млекопитающих локализовано в женских гонадах – яичниках (локализованный оогенез). В случае локализованного оогенеза ооцит может развиться без участия вспомогательных питающих клеток (солитарный тип) или при участии специальных клеток (алиментарный тип). Солитарный оогенез встречается у некоторых кишечнополостных, червей и моллюсков. Алиментарный тип оогенеза может быть фолликулярным или нутриментарным. При фолликулярном типе оогенеза растущий ооцит окружен фолликулярными (соматическими по происхождению) клетками, которые вместе образуют функциональную структуру – фолликул. Это наиболее распространенный оогенез. Он встречается у подавляющего большинства животных, к числу которых относятся все хордовые. Фолликулярные клетки не участвуют в синтезе белков желтка, все виды РНК синтезируются в самом ооците. Среди многочисленных функций фолликулярного эпителия – проведение и селекция гетеросинтетических макромолекул, защитная, регуляторная, синтез вторичных оболочек и др. При нутриментарном типе оогенеза ооцит окружен трофоцитами, связанными с ним цитоплазматическими мостиками. Трофоциты – абортивные половые клетки, т.е. имеющие общее происхождение с ооцитом. На один ооцит приходится огромное количество трофоцитов, снабжающих половую клетку РНК. В вителлогенезе трофоциты участия не принимают: желток образуется за счет поступлений высокомолекулярных веществ из вне. У млекопитающих оогенез протекает по тому же типу, что и сперматогенез, однако имеет некоторые особенности. Женская половая клетка, завершившая развитие (яйцеклетка), обладает гаплоидным ядром, сложно организованной цитоплазмой и всеми факторами, необходимыми для инициации и поддержания развития.
В оогенезе различают 3 периода: размножение, рост и созревание.
В период размножения диплоидные оогонии совершают определенное, характерное для данного вида число митотических делений, а затем переходят к стадии роста и становятся ооцитами первого порядка (ооцит 1). Особенностью оогенеза, отличающей его от спермиогенеза, является меньшая митотическая активность оогониев. У высших позвоночных пролиферация оогониев происходит только в эмбриональный период и запас половых клеток (ооцитов) в яичниках взрослых самок не пополняется (рис. 4).

Рисунок 4 – Схема строения яичника. Последовательные стадии созревания фолликулов: 1– зачатковый эпителий, 2 – первичный фолликул, 3 – многослойный фолликул, 4 – образование полости граафова пузырька, 5 – зрелый графов пузырек, 6 – атрезия фолликула, 7 – выход яйцевой клетки, 8 – желтое тело, 9 – сосуды
В период роста осуществляются два процесса: в ядре происходит профаза мейоза, а в цитоплазме растущего ооцита 1 накапливаются органеллы, питательные вещества, источники энергии, необходимые для обеспечения потребностей зародыша. Рост ооцита неравномерен. Он длится очень долго – недели, месяцы и даже годы. В нем различают два этапа: малый рост, когда ассимилируются новые вещества и ими обогащается преимущественно цитоплазма, пропорционально увеличиваются объемы ядра и цитоплазмы. В цитоплазме накапливаются РНК, белки, рибосомы, органеллы. Этот этап продолжается до середины диплотены. В ядре происходят события, связанные с коньюгацией гомологичных хромосом и кроссинговером. Приблизительно с середины диплотены ооциты 1 вступают в период большого роста или вителлогенеза, при этом увеличивается объем цитоплазмы (в сотни, тысячи раз) и она постепенно заполняется желточными включениями. Помимо желтка в этот период откладываются углеводы, липиды, рибосомы, но основное вителлогенез. Желток представляет собой смесь белков, жиров, углеводов, основной из них белок вителлогенин. По происхождению он может быть эндогенным, синтезируемым самим ооцитом, и экзогенным, который синтезируется в печени или жировом теле. У большинства животных желточные белки относятся к экзогенным. Из вителлогенина в ооците образуется вителлин, состоящий из липовителлина и фосфитина. Вителлогенин поступает в ооцит путем пиноцитоза, в цитоплазме пиноцитозные пузырьки сливаются и образуются гранулы примордиального желтка. К ним присоединяются элементы аппарата Гольджи и формируются дефинитивные желточные гранулы. Постепенно весь ооцит заполняется желточными и липидными включениями, за исключением цитоплазмы вокруг ядра и по периферии. Ооцит увеличивается в размерах в тысячи раз. Глубокие изменения в периоде роста претерпевает и ядро, что сказывается внешне в его укрупнении. Оно сильно набухает, содержимое его как бы расплывается, а хромосомы даже могут исчезнуть из пределов оптической видимости. Размеры клеток колоссально возрастают. Затем овоцит 1-го порядка вступает в период созревания, или мейоз. Здесь же совершается редукционное и эквационное деление. Процессы деления в ядре протекают совершенно, так же как и при мейозе мужского ряда, судьба же цитоплазмы иная. При редукционном делении одно ядро увлекает с собой большую часть цитоплазмы, а на долю другого остается лишь мизерная часть. Поэтому возникает одна полноценная клетка – овоцит 2-го порядка, и вторая крошечная – направительное, или полярное тельце. При втором эквационном делении несимметричное распределение цитоплазмы повторяется и опять образуется одна крупная полноценная клетка – овотида и маленькое полярное тельце. Овотида по составу ядра и функционально является вполне зрелой. Период формирования в овогенезе отсутствует. Таким образом, в овогенезе из одной овогонии возникает только одна зрелая яйцеклетка, полярные же тельца остаются недоразвитыми, рассасываются и погибают. Зрелые женские гаметы называют яйцеклетками или яйцами, а отложенные в воду икрой.
Женская половая клетка – яйцеклетка – высокоспециализированная гаплоидная клетка организма, обладающая потенциальной способностью к развитию, в отличие от сперматозоидов, крупная и неподвижная. У большинства млекопитающих и человека ее размеры достигают 100 − 200 мкм (у человека 130 мкм в диаметре, у мыши – 60, у коровы – 100, у кроликов, кошек и собак 120 − 130 мкм). Вариации размеров в основном не зависят от количества желтка. В ее цитоплазме запасено все, что необходимо для роста и развития зародыша. Если сперматозоид в ходе спермиогенеза избавляется от большей части своей цитоплазмы, то ооцит активно накапливает различные трофические компоненты и органеллы, РНК, структурные белки, предшественники синтеза ДНК и РНК, а также желток, который служит источником питания для зародыша. Количество и распределение желтка имеют важное значение для развития. Яйцеклетки животных, у которых личинки или зародыши рано переходят на активное питание, содержат мало желтка. В тех случаях, когда яйцеклетка содержит большие запасы желтка, из яйцевых оболочек выходит вполне сформированная особь. В яйцеклетках плацентарных млекопитающих, чьи эмбрионы развиваются внутриутробно и получают питательные вещества из материнского организма, объем желтка составляет менее 5 % объема яйца. Другими словами, количество желтка определяет условия и продолжительность развития зародыша. По количеству желтка различают яйцеклетки: многожелтковые (полилецитальные), маложелтковые (олиголецитальные), содержащие среднее количество желтка (мезолецитальные), безжелтковые (алецитальные). Количество желтка видоспецифично и не зависит от условий питания самки. Желток может размещаться в клетке равномерно (изо-или гомолецитальные), поэтому различают клетки телолецитальные, у которых желток преимущественно или полностью концентрируется у вегетативного полюса, и центролецитальные, у которых он собран в центральной зоне. В случае телолецитальных яйцеклеток полюс, на котором расположен желток, называется вегетативным, а противоположный ему – анимальным. У плацентарных млекопитающих они практически не содержат желтка и характеризуются как алецитальные.
На ультрамикроскопическом уровне яйцеклетка человека имеет крупное округлое ядро, в котором преобладает эухроматин, и цитоплазму с умеренным количеством митохондрий с немногочисленными ламеллярными кристами. Хорошо развита система белкового синтеза из-за высокого содержания рибо- и полисом, коротких канальцев гранулярной эндоплазматической сети. В них отсутствует центросома. В ядрах различимы хроматиновые нити и ядрышки. В цитоплазме редко и диффузно расположены немногочисленные скопления вителлиновых гранул – коллекторов трофических включений.
У высших плацентарных млекопитающих и человека женские половые клетки не существуют изолированно. За исключением ранних стадий развития (первичных половых клеток, овогоний), они находятся в тесном контакте с соматическими клетками яичника (фолликулярными эпителиоцитами и соединительнотканными клетками), которые формируют вокруг каждой половой клетки эпителиальную и соединительнотканную оболочки. Комплекс «половая клетка – соматические оболочки» именуется овариальным фолликулом. Строение овариального фолликула усложняется в процессе овогенеза.
Кроме плазматической оболочки, характерной для каждой животной клетки, яйцеклетки обычно окружены специализированными оболочками. Различают первичные, вторичные и третичные оболочки. Первичные строятся самим яйцом. Это уплотненный наружный слой его цитоплазмы. Вторичные оболочки образуются фолликулярными клетками, окружающими половую клетку. Третичные оболочки формируются из веществ, выделяемых яйцеводами во время прохождения по ним яйца. Первичная оболочка – желточная – прилегает к поверхности яйцеклетки и присуща всем животным; иногда бывает пронизана выростами яйца, выглядит радиально исчерченной и называется zona radiata. Вторичные и третичные оболочки есть не у всех яиц (рис. 5). 3 2 1

Рисунок 5 – Яйцо женщины: Яйцо (2) окружено фолликулярными клетками; видны сперматозоиды (3); некоторые проникли через барьер из фолликулярных клеток; слева видно полярное тельце (1)
ПРАКТИЧЕСКАЯ ЧАСТЬ
|
|
|
