 |


26. Метаболизм. Катаболизм и анаболизм. Схема обмена веществ.
|
|
|
|
26. Метаболизм. Катаболизм и анаболизм. Схема обмена веществ.
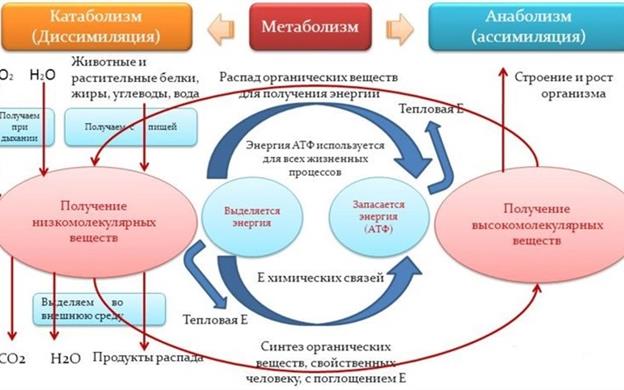
Метаболизм - это обмен веществ и превращение энергии в клетке, сложная цепь превращений веществ в организме начиная с момента их поступления из внешней среды и кончая удалением продуктов распада. В процессе обмена организм получает вещества для построения клеток и энергию для жизненных процессов. Поэтому выделяются два вида обмена: пластический и энергетический. Пластическим обменом (анаболизм или ассимиляция) — это совокупность реакций, способствующих построению клетки и обновлению ее состава. Энергетический обмен (катаболизм, диссимиляция) — совокупность реакций, обеспечивающих клетку энергией

27. Катаболизм углеводов. Переваривание углеводов. Внутриклеточный обмен углеводов. Распад и синтез гликогена. Биологическое значение обмена гликогена в печени и мышцах.
Переваривание:
Углеводы составляют около 60—70% от общей суммы калорий пищи человека, их основная масса представлена поли- и олигосахаридами. Углеводы пищи расщепляются в пищеварительном тракте до моносахаридов, которые всасываются через слизистую оболочку кишечника в кровь.
Переваривание углеводов происходит при действии амилолитических ферментов (гликозидаз), катализирующих гидролиз гликозидных связей. К ним относятся слюнная и панкреатическая а-амилаза. В амилопектине значительная часть молекулы остается нерасщепленной. Такой частично переваренный амилопектин называется декстрином, в тонком кишечнике он расщепляется ферментом амило-а-(1— 6)-глюкозидазой. Под действием этого фермента образуются ди- и трисахариды, атакуемые мальтазой. Основным местом переваривания углеводов является тонкий кишечник, где на них действует а-амилаза поджелудочной железы и активные дисахарозидазы. Моносахариды, образующиеся в результате последовательного действия амилолитических ферментов, с высокой эффективностью, но совершенно разными скоростями всасываются в кровь. Манноза и пентозы проникают через эпителий кишечника только путем облегченной диффузии с участием специальных переносчиков. Галактоза и глюкоза кроме этого пути могут транспортироваться против градиента их концентрации по механизму вторичного активного транспорта. Поступление глюкозы из крови в клетки осуществляется в направлении падения ее градиента, так как в цитозоле большинства животных клеток концентрация свободной глюкозы очень низка, тогда как концентрация в плазме крови близка к 5 ммоль/л. Однако только в клетки печени и мозга транспорт глюкозы может осуществляться по механизму пассивной диффузии, и скорость поступления регулируется ее концентрацией в крови. Во всех других тканях скорость транспорта глюкозы осуществляется по механизму облегченной диффузии, который стимулируется инсулином.
|
|
|
Внутриклеточный обмен углеводов.
Общая характеристика:
Во всех клетках, способных метаболизировать глюкозу, первой реакцией является ее фосфорилирование до глюкозо-6-фосфата. Реакция катализируется ферментом гексокиназой, а донором фосфорильной группы является молекула АТФ:

Эта реакция практически необратима, Δ G= -16, 74 кДж/моль. Гексокиназа, присутствующая во всех тканях, за исключением паренхимы печени, имеет высокое средство к глюкозе, а также способна фосфорилировать и другие гексозы, но значительно с меньшей скоростью. В клетках печени эту функцию выполняет глюкокиназа, активность которой зависит от питания. Глюкокиназа специфична к глюкозе и эффективно функционирует только при высокой концентрации в крови глюкозы. Важным свойством гексокиназы является ее ингибирование продуктом реакции глюкозо-6- фосфатом по аллостерическому механизму. Фосфорилированная глюкоза не способна проходить через цитоплазматическую мембрану и оказывается «запертой» в клетке. Таким образом, глюкозо-6-фосфат является центральным метаболитом углеводного обмена и занимает важное положение в интеграции ряда метаболических путей (гликолиз, глюконеогенез, пентозофосфатный путь, гликогенолиз). Обратный процесс дефосфорилирования глюкозы идет только в трех тканях, клетки которых способны транспортировать глюкозу в кровь, а именно ткани печени, эпителия почечных канальцев и тонкого кишечника. Это становится возможным благодаря действию гидролитического фермента глюкозо-6-фосфатазы, который катализирует реакцию. О регуляции активности этого фермента до сих пор известно мало, а следовательно, неясно, какие факторы предотвращают непрерывный цикл фосфорилирования и дефосфорилирования глюкозы.
|
|
|

|
|
|
