 |


Рассмотрим подробнее, что же происходит в нашем организме, когда в него проникают дрожжи.
|
|
|
|
Термофильные дрожжи и их негативное влияние на здоровье.
Итак, повторим: дрожжи-сахаромицеты (термофильные дрожжи), различные расы, которых употребляются в спиртовой промышленности, пивоварении и хлебопечении, в диком состоянии в природе не встречаются, то есть это создание рук человеческих. Они относятся по морфологическим признакам к простейшим сумчатым грибам и микроорганизмам.
Сахаромицеты, к несчастью, являются более совершенными, чем тканевые клетки, независимы от температуры, рН среды, содержания воздуха. Даже при разрушенной лизоцимом слюны оболочки клетки они продолжают жить.
Производство пекарских дрожжей основано на размножении их в жидких питательных средах, приготовляемых из мелассы (отходов от производства сахара).
Технология чудовищная, антиприродная. Мелассу разбавляют водой, обрабатывают хлорной известью, подкисляют серной кислотой и т. д. Странные методы, надо признать, используются для приготовления пищевого продукта, к тому же, если учесть, что в природе существуют естественные дрожжи, хмелевые, например, солод и т. д.
А теперь посмотрим, какую "медвежью услугу" оказывают термофильные дрожжи нашему организму.
Достоин внимания опыт французского учёного Этьена Вольфа. Он в течение 37 месяцев культивировал злокачественную опухоль желудка в пробирке с раствором, в котором находился экстракт ферментирующих дрожжей. В это же время в течение 16 месяцев культивировалась в таких же условиях, вне связи с живой тканью, опухоль кишечника.
В результате эксперимента выяснилось, что в таком растворе размер опухоли удваивался и утраивался в течении одной недели. Но как только из раствора удалялся экстракт, опухоль погибала. Отсюда был сделан вывод, что в экстракте дрожжей содержится вещество, стимулирующее рост раковых опухолей.
|
|
|
Учёные Канады и Англии установили убивающую способность дрожжей.
Клетки-килеры, клетки-убийцы дрожжей убивают чувствительные, менее защищенные клетки организма путём выделения в них ядовитых белков малого молекулярного веса. Токсичный белок действует на плазменные мембраны, увеличивая их проницаемость для патогенных микроорганизмов и вирусов. Дрожжи попадают вначале в клетки пищеварительного тракта, а потом уже в кровяное русло.
Таким образом, они становятся тем "троянским конем", с помощью которого неприятель попадает в наш организм и способствует подрыву его здоровья.
Термофильные дрожжи настолько реактивны и живучи, что при 3-4 кратном использовании их активность только возрастает. Известно, что при выпечке хлеба дрожжи не уничтожаются, а сохраняются в капсулах из клейковины. Попадая в организм, они начинают свою разрушительную деятельность.
Сейчас уже хорошо известно специалистам, что при размножении дрожжей формируются аскоспоры, которые, оказываясь в нашем пищеварительном тракте, а затем, попадая в кровеносное русло, разрушают мембраны клеток, способствуя онкологическим заболеваниям.
Современный человек употребляет много пищи, но наедается с трудом. Почему?
Да потому, что спиртовое брожение, осуществляемое дрожжами, без доступа кислорода, является процессом неэкономичным, расточительным с биологической точки зрения, так как из одной молекулы сахара выделяется всего 28 ккал, в то время как при широком доступе кислорода освобождается 674 ккал.
Дрожжи размножаются в условиях организма в геометрической прогрессии и позволяют патогенной микрофлоре активно жить и размножаться, угнетая нормальную микрофлору, благодаря которой в кишечнике могут вырабатываться при правильном питании и витамины группы В, и незаменимые аминокислоты.
|
|
|
По заключению академика Ф. Углова дрожжевые компоненты, попадающие в пищу, провоцируют выработку в организме дополнительного этанола. Не исключено, что это является одним из факторов, сокращающих человеческую жизнь. Развивается ацидоз, которому способствует выделяемый при спиртовом брожении уксусный альдегид и уксусная кислота, являющиеся конечным продуктом превращения спирта.
В период прикармливания ребёнка кефиром к этанолу грудного молока добавляется кефирный этанол. В пересчёте на взрослый мужской эквивалент это равносильно ежесуточному потреблению водки от рюмки до стакана и более. Вот так происходит процесс алкоголизации России. Наша страна оказалась единственной в мире из 212 стран планеты с крупномасштабным кормлением детей слабоалкогольным кефиром. Задумайтесь, кому это нужно?
Направленный против здоровья человека союз дрожжей и молочнокислых бактерий приводят организм в конечном итоге к некомпенсированной стадии ацидоза.
Чрезвычайно интересно исследование В. М. Дильмана, доказывающего, что онкоген газ содержит дрожжи, А. Г. Качужный и А. А. Болдырев своими исследованиями подтвердили сообщение Этена Вольфа о том, что дрожжевой хлеб стимулирует рост опухоли. В. И. Гринев обращает внимание на то, что в США, Швеции и других странах бездрожжевой хлеб стал обычным явлением и рекомендуется как одно из средств профилактики и лечения онкозаболеваний.
Рассмотрим подробнее, что же происходит в нашем организме, когда в него проникают дрожжи.
Грубейшим образом нарушается деятельность всех органов пищеварения при брожении, особенно вызванном дрожжами. Брожение сопровождается гниением, развивается микробная флора, травмируется щеточная кайма, патогенные микроорганизмы с лёгкостью проникают через стенку кишечника и попадают в ток крови. Замедляется эвакуация токсических масс из организма, образуются газовые карманы, где застаиваются каловые камни. Постепенно они врастают в слизистые и подслизистые слои кишечника. Продолжает нарастать интоксикация продуктами жизнедеятельности бактерий, бактериемия (когда они «осеменяют» нашу кровь). Секрет органов пищеварения утрачивает защитную функцию и снижает пищеварительную.
|
|
|
Недостаточно усваиваются и синтезируются витамины, не усваиваются в должной мере микроэлементы и главнейший из них - кальций, происходит сильная утечка кальция с целью нейтрализации разрушительного действия избыточных кислот, которые появляются в результате аэробного брожения.
Использование в пищу дрожжевых продуктов способствует не только канцерогенезу, то есть образованию опухолей, но и запорам, усугубляющим канцерогенную ситуацию, образованию сгустков песка, камней в желчном пузыре, печени, поджелудочной железе; жировой инфильтрации органов или наоборот - дистрофическим явлениям и в конечном итоге ведёт к патологическим изменениям важнейших органов.
Серьёзным сигналом о запущенном ацидозе является увеличение холестерина в крови сверх нормы. Истощение буферной системы крови приводит к тому, что свободные избыточные кислоты травмируют внутреннее покрытие сосудов. Холестерин в виде шпаклевочного материала начинает использоваться для залатывания дефектов.
При брожении, которое вызывают термофильные дрожжи, возникают не только негативные физиологические изменения, но даже анатомические. В норме сердце и лёгкие и нижележащие органы - желудок и печень, а также поджелудочная железа получают мощный массирующий энергетический стимул от диафрагмы, которая является главной дыхательной мышцей, взлетающей до 4-го и 5-го межреберья.
При дрожжевом брожении диафрагма не выполняет колебательных движений, занимает вынужденную позицию, сердце располагается горизонтально (в положении относительного покоя), оно часто ротировано (то есть, развернуто относительно своей оси), нижние доли лёгких сдавлены, все органы пищеварения зажаты чрезвычайно раздутым газами деформированным кишечником, часто желчный пузырь покидает своё ложе, изменяя даже форму.
В норме диафрагма, совершая колебательные движения, способствует созданию присасываюшего давления в грудной клетке, которое притягивает кровь от нижних и верхних конечностей и головы на очищение в лёгкие.
|
|
|
При ограничении её экскурса, этого не происходит. Всё это вместе способствует нарастанию застойных явлений в членах нижних конечностей, малого таза и головы и в итоге - варикозному расширению вен, тромбообразованию, трофическим язвам и дальнейшему снижению иммунитета. В результате человек превращается и плантацию для произрастания вирусов, грибков, бактерий, риккетсий (клещей).
Когда сотрудники фирмы "Виватон" работали в Институте патологии кровообращения в Новосибирске, то они получили убедительные свидетельства от академика Мешалкина и профессора Литасовой о том, какое негативное опосредованное влияние оказывает дрожжевое брожение на деятельность сердца.
«ХЛЕБОПЕКАРНЫЕ ДРОЖЖИ - сделано по ГОСТу...»

Так из чего же изготавливаются хлебопекарные дрожжи, которые мы каждый день употребляем в составе различных хлебобулочных изделий?
Для выработки хлебопекарных дрожжей (по ГОСТ 171-81) используется следующее основное и вспомогательное сырьё:
- свекловичная мелласа с рН среды 6,5 до 8,5 с массовой долей сахарозы не менее 43,0% с массовой долей суммы сбраживаемых сахаров не менее 44,0% по ОСТ 18-395;
- сульфат аммония по ГОСТ 3769;
- сульфат аммония технический, полученный при производстве сернистого ангидрида;
- аммоний сернокислыйочищенный по ГОСТ 10873;
- аммоний гидроортофосфат марки А по НТД;
- аммиак водный технический марки Б (для промышленности) по ГОСТ 9;
- карбамид по ГОСТ 2081;
- диаммонийфосфат технический (для пищевой промышленности) по ГОСТ 8515;
- вода питьевая по ГОСТ 2874*;
- кислота ортофосфорная термическая по ГОСТ 10678;
- калий углекислый технический (поташ) по ГОСТ 10690 первого сорта;
- калий хлористый по ГОСТ 4568 марки;
- калий хлористый технический по НТД;
- магний сернокислый 7-водный по ГОСТ 4523;
- магний хлористый технический (бишофит) по ГОСТ 7759;
- эпсомит;
- порошок магнезитовый каустический по ГОСТ 1216;
- экстракт кукурузный сгущенный;
- дестиобиотин ЦТД;
- кислота серная техническая по ГОСТ 2184 (контактная улучшенная марок А и Б) или аккумуляторная по ГОСТ 667;
- мальц-экстракт;
- солод пивоваренный ячменный;
- сильвинит;
- микроудобрение для сельского хозяйства южных районов;
- мел химически осажденный по ГОСТ 8253;
- крахмал картофельный по ГОСТ 7699;
- соль поваренная пищевая по ГОСТ 13830*;
- бельтинг хлопчатобумажный фильтровальный по ГОСТ 332;
- пеногасители;
- кислота олеиновая; техническая (олеин) по ГОСТ 7580, марок Б14 и Б16;
- кислота олеиновая техническая (олеин) марки «О» или марки ОМ;
- дистиллированные жирные кислоты подсолнечного и соевого масел;
- масло хлопковое рафинированное по ГОСТ 1128;
- концентрат пекарский фосфатидный;
- масло подсолнечное по ГОСТ 1129;
- дезинфицирующие вещества;
- известь хлорная по ГОСТ 1692;
- известь строительная по ГОСТ 9179;
- известь белильная (термостойкая);
- натр едкий технический по ГОСТ 1625;
- кислота молочная пищевая по ГОСТ 490;
- кислота борная по ГОСТ 9656;
- водорода перекись по ГОСТ 177;
- фурацилин;
- фуразолидон;
- сульфонол НП-3;
- катапин (бактерицидный);
- моющее жидкое средство «Прогресс»;
- калий маргонцовокислый технический по ГОСТ 5777;
- кислота соляная синтетическая техническая по ГОСТ 857;
- кальция пантотенат по ФС 42-2530;
- кальция пантотенат рацемический для животноводства по НТД;
- кислота соляная техническая по НТД;
- кислота соляная из хлористого водорода-ректификата марки Б по НТД.
|
|
|
Таким образом, из более чем полусотни составляющих в пищу без вреда для здоровья можно употреблять лишь около 10!
Как видно из официального государственного документа, для производства хлебопекарных дрожжей используется 36 видов основного и 20 видов вспомогательного сырья, абсолютное большинство которого никак не назовешь пищевым. С помощью микроудобрений для сельского хозяйства южных районов и прочих химикалий дрожжи насыщаются тяжелыми металлами (медь, цинк, молибден, кобальт, магний и пр.) и иными не всегда полезными нашей плоти химическими элементами (фосфор, калий, азот и т.д.). Их роль в процессе дрожжевого брожения ни в каких справочниках не раскрывается...
Сейчас ученые пришли к выводу о том, что дрожжевой хлеб является одной из причин возникновения рака. Но до сих пор, технология производства дрожжевого хлеба не изменилась. И здесь всё ясно, если хочешь жить в здоровом теле - перестань есть дрожжевой хлеб.
Кто предупреждён - тот вооружён! Берегите своё здоровье и здоровье своих близких!
«Дрожжи помнят о несчастной любви.»

Особый белок, накапливающийся в клетках дрожжей, не даёт им забыть о неудачных попытках полового процесса, и в дальнейшем, чтобы «раскрутить» такие клетки на половое размножение, их потенциальным партнёрам придётся потратить изрядное количество феромонов.
У дрожжей, кроме бесполого размножения, когда дочерняя клетка отпочковывается от материнской, есть и половой процесс — когда две клетки сливаются вместе, объединяя два генома в один. Сливаться могут не любые клетки, а гаплоидные (то есть не с двумя, а с одним набором хромосом), и притом разные по типу спаривания — грубо говоря, «разнополые» (хотя говорить о половой принадлежности дрожжей вряд ли стоит). Такие клетки выделяют особые химические вещества, которые может распознать клетка-партнёр. Если клетки нашли друг друга, они протягивают специальные выросты, и если выросты соприкоснулись, произойдёт слияние, за которым последует ряд довольно сложных процессов.
Но бывает так, что слияния не происходит, и все химические «признания в любви» и протягивание выростов оканчиваются неудачей. В этом случае, как пишут в журнале Cell исследователи из Швейцарской высшей технической школы Цюриха, в дрожжах накапливается неактивная форма белка Whi3. Этот белок и так присутствует в клетке, но стоит дрожжам испытать любовную неудачу, Whi3 изменяет пространственную конформацию, причём изменённые молекулы заражают ею другие молекулы Whi3.
В итоге неактивный Whi3 образует агрегаты, которые сама клетка если и может разрушить, то с огромным трудом. Но самое интересное то, как такие агрегаты влияют на половое размножение: в следующий раз потенциальному партнёру придётся потратить гораздо больше феромонов, чтобы заинтересовать клетку, которая перенесла неудачную любовь. Если таких веществ окажется недостаточно, клетка просто не будет обращать внимание на призывы и продолжит размножаться бесполым способом, то есть простым почкованием.
Фактически у клетки формируется память о «несчастных любовях», и чем старше становится клетка, тем сложнее ей найти партнёра: число разочарований, знаете ли, со временем растёт, и белок накапливается, отбивая у дрожжей всякое желание «интима». Лишь в очень редких случаях клетке удаётся очистить свою память. Стоит также заметить, что к дочерним клеткам, образовавшимся в результате почкования, «груз памяти» не переходит, поэтому им не возбраняется участие в половом процессе.
Причину накопления такой памяти учёные объясняют следующим образом. Дрожжам приходится выбирать, размножаться быстро бесполым образом, но при этом снижать генетическую вариабельность популяции, или размножаться половым способом, увеличивая генетическое разнообразие потомства, но тратя на это много энергии. Когда клетка дрожжей чувствует феромоновый «любовный призыв», она отключает бесполое деление, готовясь к половой встрече. Но потенциальный партнёр может быть обманщиком, который таким образом тормозит размножение конкурентов. Поэтому клетка с каждым таким обманом поднимает ставку: потенциальный партнёр должен действительно доказать, что он намерен вступить в половое размножение, и доказать это он должен повышенным уровнем феромонов.
Любопытно, что нечто похожее происходит у дрозофил: когда самка по какой-то причине отвергает самца, в его синапсах появляются белковые отложения, которые, возможно, влияют на его дальнейшее брачное поведение. Не исключено, что и у высших животных есть подобный способ запоминания. Правда, в случае человеческих мозгов словосочетание «белковые отложения» вызывает неприятные ассоциации с болезнью Альцгеймера и прочими нейродегенеративными недугами. Было бы удивительно вдруг обнаружить, что нейродегенеративные процессы — это всего лишь вышедшие из-под контроля процессы памяти...
| |||||||
|
Та же история, что и с НИИ мясомолочной промышленности, и большей части фармацевтики. Пытаются защищаться и оправдать вредительство.
| Эволюция полового размножения у дрожжей: надстройка меняется, базис остается |
25.02.14 | Генетика, Биология развития, Эволюция, Александр Марков | Комментировать
|
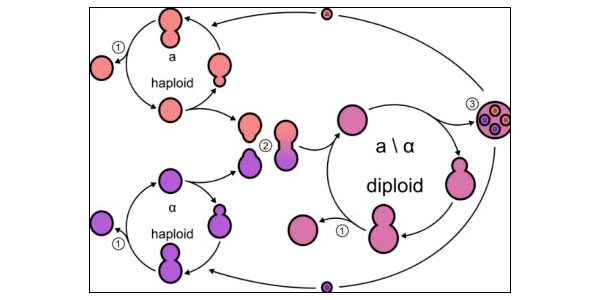 Рис. 1. Схема жизненного цикла пекарских дрожжей Saccharomyces cerevisae. 1 — почкование, 2 — конъюгация (спаривание), 3 — образование четырех гаплоидных спор из диплоидной клетки в результате мейоза. Схема с сайта www.wynboer.co.za
Рис. 1. Схема жизненного цикла пекарских дрожжей Saccharomyces cerevisae. 1 — почкование, 2 — конъюгация (спаривание), 3 — образование четырех гаплоидных спор из диплоидной клетки в результате мейоза. Схема с сайта www.wynboer.co.za
Американские биологи обнаружили, что генные сети, регулирующие половое размножение, по-разному устроены у разных видов дрожжей. У обычных пекарских дрожжей и ряда других видов спаривание (слияние двух гаплоидных клеток в одну диплоидную) и мейоз (образование гаплоидных спор из диплоидной клетки) разнесены во времени, а в жизненном цикле преобладает диплоидная фаза. У других видов мейоз происходит вскоре после спаривания, и поэтому в жизненном цикле преобладает гаплоидная фаза. Как выяснилось, в первом случае спаривание и мейоз управляются двумя независимыми генетическими программами, а во втором эти программы слиты воедино, причем белки-регуляторы, необходимые для спаривания, стали необходимыми также и для мейоза, и наоборот. По-видимому, слияние управляющих программ происходило независимо в разных группах дрожжей, когда им становилось выгодно проводить большую часть жизни в гаплоидном состоянии.
У организмов, размножающихся половым путем, в жизненном цикле обязательно присутствуют два особых события – оплодотворение, в результате которого два гаплоидных генома объединяются в одной клетке, и редукционное деление (мейоз), в ходе которого из диплоидной клетки получаются гаплоидные.
Иногда эти события тесно связаны и следуют одно за другим, а иногда — разнесены во времени и происходят более или менее независимо. Первый вариант характерен, например, для животных (у которых оплодотворение следует за мейозом), а также для многих одноклеточных, проводящих большую часть жизни в гаплоидном состоянии (в этом случае, наоборот, мейоз происходит сразу после оплодотворения).
Пример второго варианта — пекарские дрожжи Saccharomyces cerevisae, классический лабораторный объект. Диплоидные клетки S. cerevisae могут долго размножаться почкованием, производя дочерние диплоидные клетки. В определенных условиях (например, при голодании) диплоидная клетка претерпевает мейоз и производит четыре гаплоидные споры. Из спор вырастают гаплоидные дрожжевые клетки, которые тоже могут размножаться почкованием, однако они, в отличие от диплоидных клеток, способны еще и к спариванию (конъюгации). Гаплоидные дрожжи делятся на два пола (a и α), причем спариваться могут только разнополые клетки. В результате спаривания получается диплоидная клетка, и цикл продолжается (рис. 1). Таким образом, у S. cerevisae между мейозом и оплодотворением могут быть долгие периоды бесполого размножения, а сами эти два события друг с другом напрямую не связаны и регулируются разными генами.
По-другому устроен жизненный цикл родственника пекарских дрожжей, патогенного гриба Candida lusitaniae. У этого вида большая часть жизни проходит в гаплоидной фазе, а образовавшаяся в результате конъюгации диплоидная клетка, как правило, не почкуется (или почкуется недолго) и вскоре приступает к мейозу.
Микробиологи из Брауновского университета (США) решили разобраться в причинах этого различия. Они сравнили экспрессию генов S. cerevisae и C. lusitaniae во время спаривания и мейоза. У S. cerevisae наборы генов, отвечающих за эти два процесса, четко различаются. Однако у C. lusitaniae, как выяснилось, разделение труда между двумя группами генов выражено намного слабее.
Во время спаривания у C. lusitaniae активируются не только гены, которые у пекарских дрожжей отвечают за спаривание (включая гены MAPK-каскада, обеспечивающего реакцию клетки на половые феромоны противоположного пола), но и некоторые гены, которые у S. cerevisae необходимы для мейоза, но в спаривании не участвуют. К числу таких «генов мейоза», активирующихся у C. lusitaniae во время спаривания, относятся SPO11, REC8 и IME2.
В диплоидных клетках C. lusitaniae, собравшихся приступить к мейозу, резко (более чем в четыре раза) возрастает экспрессия 618 генов. Самое интересное, что многие из этих генов у S. cerevisae обслуживают исключительно процесс спаривания, а в мейозе не участвуют (у S. cerevisae при мейозе возрастает экспрессия меньшего числа генов — 480). В частности, многие гены, у пекарских дрожжей и других грибов отвечающие за реагирование на половые феромоны, у C. lusitaniae почему-то активируются не только при подготовке к спариванию, но и в ходе мейоза. С другой стороны, некоторые гены, активирующиеся у S. cerevisae при спаривании или мейозе, у C. lusitaniae в этих ситуациях не активируются. Ключевой регулятор мейоза IME1 вообще отсутствует у C. lusitaniae.
Полученные факты позволяют заключить, что генные сети, регулирующие спаривание и мейоз у двух видов, существенно различаются. Чтобы выяснить, имеют ли эти различия функциональное значение, авторы приступили к изучению работы отдельных генов.
Ген IME2 (inducer of meiosis 2) является ключевым регулятором мейоза у S. cerevisae. Как и следовало ожидать, удаление этого гена у C. lusitaniae лишило диплоидные клетки способности к мейозу. Но это не всё: гаплоидные клетки C. lusitaniae без гена IME2 оказались неспособны к спариванию. Между тем у S. cerevisae ген IME2 на спаривание не влияет. Половые феромоны необходимы пекарским дрожжам для регуляции спаривания, но не мейоза. Оказалось, что C. lusitaniae активно производят феромоны и в ходе мейоза, и при спаривании. Ген STE12 кодирует транскрипционный фактор, играющий ключевую роль в регуляции полового поведения у S. cerevisae и многих других грибов. Его удаление у C. lusitaniae лишило клетки не только способности к спариванию (этот результат был ожидаем), но и способности к мейозу, при том что у S. cerevisae этот ген в мейозе не задействован.
Вся совокупность данных говорит о том, что у S. cerevisae есть две более или менее самостоятельные генетические программы, одна из которых регулирует половое поведение (спаривание), а другая — мейоз. У родственного вида C. lusitaniae эти две программы фактически слились в одну.
Авторы проверили, влияет ли удаление гена STE12 на мейоз еще у трех близких видов дрожжей: Kluyveromyces lactis, Pichia pastoris и Yarrowia lipolytica. Во всех трех случаях никакого влияния выявлено не было (как и у S. cerevisae). Это значит, что разделенность программ спаривания и мейоза, по-видимому, является исходным (предковым) признаком данной группы грибов, а их слияние у C. lusitaniae — признак новоприобретенный (рис. 2).
Возможно, ключом к слиянию генетических программ послужила утрата транкрипционного фактора α2. Об этом белке известно, что он не позволяет некоторым генам, активным в гаплоидных клетках (в том числе генам MAPK-каскада, отвечающего за реагирование на половые феромоны), работать также и в диплоидной фазе жизненного цикла. Предки C. lusitaniae утратили α2, что дало возможность «генам спаривания» работать в диплоидных клетках. Это и позволило некоторым из них взять на себя новую функцию регуляторов мейоза.
Слияние программ спаривания и мейоза не является уникальной особенностью C. lusitaniae. Такое же явление обнаружено у дальнего родственника рассматриваемой группы, Schizosaccharomyces pombe, чьи предки отделились от предков других дрожжей более 330 млн лет назад. У этого вида, как и у C. lusitaniae, транскрипционные факторы MAPK-каскада, необходимые для спаривания, необходимы также и для мейоза.
|
 Рис. 2. Вверху (a) — схема регуляции спаривания (Mating) и мейоза (Meiosis) у некоторых видов дрожжей. У Schizosaccharomyces pombe и Candida lusitaniae оба процесса регулируются совместно, с участием половых феромонов и белков MAPK-каскада, и индуцируются голоданием. У Saccharomyces cerevisae два процесса регулируются по отдельности, а половые феромоны нужны только для спаривания. Внизу (b) — эволюционное дерево, показывающее, что совместная регуляция спаривания и мейоза возникла у S. pombe и C. lusitaniae независимо. Справа отмечены галочками виды, у которых транскрипционные факторы Ste11 и Ste12 необходимы не только для спаривания, но и для мейоза; крестиками — те, кому они нужны только для спаривания. Рисунок из обсуждаемой статьи в Nature
Рис. 2. Вверху (a) — схема регуляции спаривания (Mating) и мейоза (Meiosis) у некоторых видов дрожжей. У Schizosaccharomyces pombe и Candida lusitaniae оба процесса регулируются совместно, с участием половых феромонов и белков MAPK-каскада, и индуцируются голоданием. У Saccharomyces cerevisae два процесса регулируются по отдельности, а половые феромоны нужны только для спаривания. Внизу (b) — эволюционное дерево, показывающее, что совместная регуляция спаривания и мейоза возникла у S. pombe и C. lusitaniae независимо. Справа отмечены галочками виды, у которых транскрипционные факторы Ste11 и Ste12 необходимы не только для спаривания, но и для мейоза; крестиками — те, кому они нужны только для спаривания. Рисунок из обсуждаемой статьи в Nature
По-видимому, слияние генетических программ спаривания и мейоза происходило в эволюции грибов неоднократно. По мнению авторов, это могло быть связано с тем, что некоторым видам почему-то становилось выгодно большую часть жизненного цикла проводить в гаплоидной фазе (хотя в чем именно состоят преимущества и недостатки гаплоидности и диплоидности — вопрос дискуссионный, см.: S. P. Otto, A. C. Gerstein, 2008. The evolution of haploidy and diploidy). Единая регуляция спаривания и мейоза у C. lusitaniae и S. pombe приводит к тому, что мейоз у них обычно происходит вскоре после образования диплоидной клетки, и поэтому большую часть жизни эти грибы проводят в гаплоидном состоянии. Напротив, у S. cerevisae и других видов с разделенными программами спаривания и мейоза в жизненном цикле преобладает диплоидная фаза.
Исследование еще раз показало, что в ходе эволюции может происходить радикальная перестройка (rewiring) генных сетей, регулирующих развитие того или иного признака, хотя сам признак при этом не меняется. Другой пример — огромное разнообразие способов детерминации пола у разных организмов, хотя в итоге получается одно и то же: часть особей становится самцами, часть — самками (впрочем, к дрожжам с их «полами» а и α это как раз не относится).
Источник: Racquel Kim Sherwood, Christine M. Scaduto, Sandra E. Torres & Richard J. Bennett. Convergent evolution of a fused sexual cycle promotes the haploid lifestyle // Nature. 2014. V. 506. P. 386–390.
Про половое размножение дрожжей см. также:
Дрожжи занимаются сексом не от хорошей жизни, «Элементы», 18.04.2012.
Про устройство MAPK-каскада (обеспечивающего реакцию дрожжей на половые феромоны противоположного пола) см. также:
Перекомбинирование фрагментов белковых молекул — быстрый способ создания новых признаков, «Элементы», 20.04.2010.
 Рис. 1. Жизненный цикл дрожжей Saccharomyces cerevisae. Изображение с сайта upload.wikimedia.org
Рис. 1. Жизненный цикл дрожжей Saccharomyces cerevisae. Изображение с сайта upload.wikimedia.org
