 |


Глава 3. Анаболизм и катаболизм липидов
|
|
|
|
Липиды – это разнообразная по строению группа органических соединений, общим свойством которых является гидрофобность. В отличие от представителей других классов липиды не могут полимеризоваться, а усложнение их осуществляется за счет присоединения самых разных по природе веществ. Простые однокомпонентные липиды – ВЖК, высшие спирты, в том числе сфингозин, ХС, не способны к гидролизу. Простые двукомпонентные липиды - сложные эфиры, то есть продукты взаимодействия ВЖК и различных спиртов (с глицерином - ТАГ, с высшими ациклическими спиртами - воска, с ХС - его эфиры). Исключением служат церамиды, являющиеся амидами ВЖК и аминоспирта сфингозина. В состав сложных липидов входят полярные компоненты (фосфорилированные азотистые основания в ФЛ, моносахариды в гликолипиды, полипептиды в ЛП), что делает их амфифилами.
Функции липидов: ТАГ – компактная и энергоемкая форма хранения энергии в адипоцитах жировой ткани, выполняющей теплоизолирующую и механическую защитные функции. Входит в состав билипидного слоя всех видов мембран, включающего различные фосфо-, гликолипиды и ХС. В коже данный стероид преобразуется в холекальциферол (витамин D3); в коре надпочечников и в половых железах из него синтезируются соответствующие гормоны; в печени – желчные кислоты (Рис. 3.10).
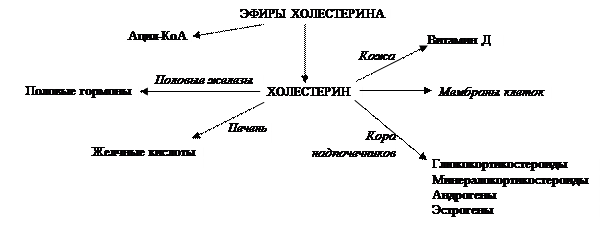
Рис. 3.10. Пути использования эфиров холестерина в клетке
Около 50% ВЖК фосфолипидов мембран являются полиненасыщенными, что увеличивает текучесть и проницаемость. Такие ВЖК очень чувствительны к действию различных радикалов, в первую очередь, активных форм кислорода (АФК): супероксида аниона О2.- , пероксида водорода, радикала гидроксила и других, которые индуцируют перекисное окисление липидов (ПОЛ), чрезмерная активация которого сопровождает или служит причиной многих патологических состояний (Рис. 3.11). В физиологических условиях эти процессы участвуют в самообновлении и репарации мембран, в синтезе БАВ. Кроме того, АФК являются фактором бактерицидности фагоцитирующих клеток.
|
|
|
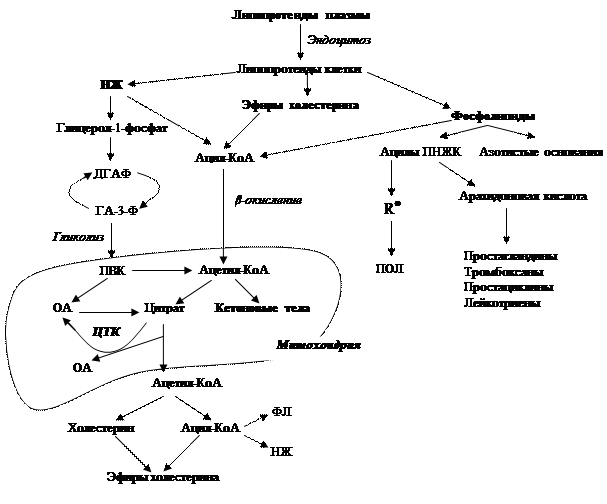
Рис. 3.11. Взаимосвязь метаболизма липидов с другими видами обменов
Ряд ПНЖК (арахидоновая и эйкозапентаеновая кислоты) участвуют в образовании простагландинов, лейкотриенов, тромбоксанов. Некоторые ПНЖК (линолевая, α-линоленовая, арахидоновая, эйкозапентаеновая и докозагексаеновая) не синтезируются в организме человека и относятся к незаменимым (эссенциальным), поэтому их объединяют под термином витамин F.
Катаболическая фаза для большинства липидов также складывается из трёх стадий. Если молекула липида состоит из двух и более компонентов, то она гидролизуется; затем продукты подвергаются специфическому распаду. Высвобождающийся глицерол фосфорилируется и окисляется до дигидроксиацетонфосфата, который вступает в гликолиз, повторяя судьбу глюкозо-6-фосфата. ВЖК, точнее их активные формы (ацил-КоА), попадая в митохондрии клеток, служат субстратами β-окисления, конечным продуктом которого является ацетил-КоА, сгорающий в цикле трикарбоновых кислот (Рис. 3.11).
Как известно, основными энергоисточниками в клетках служат глюкоза и ВЖК, но последние для своего полного распада (до углекислого газа и воды) требуют больших количеств кислорода, что, естественно, затрудняет этот процесс (отсюда понятно, почему пополнев, трудно похудеть). С энергетической целью тканями (мышечной, нервной и др.) могут использоваться кетоновые тела (ацетоацетат, β-гидроксибутират). В физиологических условиях они образуются в митохондриях гепатоцитов из ацетил-КоА (Рис. 3.11).
Основные поставщики ацетил-КоА – глюкоза и ВЖК, а ОА образуется, в первую очередь, из глюкозы. Когда нарушается утилизация глюкозы (при сахарном диабете), уменьшается синтез ОА, приводящий к снижению синтеза цитрата и при наличии высокого уровня ацетил-КоА (усиленный катаболизм ВЖК), последний конденсируется в ацетоацетат (Рис. 3.12).
|
|
|

Норма; при замедлении окисления глюкозы
Рис. 3.12. Пути использования ацетил-КоА в митохондриях печени в зависимости от скорости распада глюкозы
Следует отметить, что субстратом в образовании ХС и ВЖК тоже служит ацетил-КоА, который используется на эти цели тогда, когда угнетается его распад в ЦТК. Подобная ситуация возникает в тех условиях, если результате работы цикла Кребса и связанных с ним биологического окисления и окислительного фосфорилирования генерируется много молекул АТФ. Избыток последних ингибирует дальнейшее преобразование цитрата в ЦТК, он выходит из митохондрий, распадается на исходные составные части. При этом высвобождается ацетил-КоА, который, конденсируясь, и дает или ВЖК, или холестерин (Рис. 3.12, 3.13). Особую роль в синтезе этих соединений играет НАДФН+Н+, источником которого служит только ПФП.
Мало того, в жировой ткани накопление ТАГ определяется достаточным количеством моносахаридов (Рис. 3.13). Как видно из схемы, оба компонента нейтрального жира: и ВЖК, и глицерол-1-фосфат - образуются из фосфорилированной глюкозы. Если в липоцит и попадёт глицерол, он не способен участвовать в синтезе ТАГ-ов, так как в этих клетках отсутствует глицеролкиназа – фермент, активирующий данный спирт, без чего последний не может вступать в реакции. Цитоплазматический ацетил-КоА используется в реакциях ацетилирования с образованием ацетилхолина, ацетилглюкозамина и производных моносахаридов – составных частей гетерополисахаридов.

Рис. 3.13. Схема синтеза нейтральных жиров в адипоцитах
Особая роль в жизнедеятельности организма принадлежит ПОЛ. Его индукторами служат АФК, которые могут генерироваться в небольших количествах в физиологических условиях. При гипероксии, гипоксии, действии различных лучей (рентген,- ультрафиолетовых, инфракрасных и т.д.), токсинов и других факторов уровень свободных радикалов будет расти. Активность липопероксидации подавляется веществами, которые имеются в норме в клетках и в плазме крови. Это энзимы: глутатионпероксидаза (селен-содержащий энзим), глутатионредуктаза, каталаза, супероксиддисмутаза (СОД), а также соединения неферментативной природы (каротины, витамины А, Е, С, Р, рибофлавин, глутатион, цистеин и др.), обладающие способностью обезвреживать АФК и являющиеся ловушками радикалов.
|
|
|
В условиях дефицита факторов АРЗ и/или избыточной генерации свободных радикалов, последние начинают воздействовать на ВЖК фосфолипидов биомембран и на белки, нуклеиновые кислоты, углеводы, приводя к деструкции мембран и, в конечном итоге, гибели клеток. Этот процесс неспецифичен и является важным звеном патогенеза многих заболеваний (атеросклероза, панкреатита, ревматоидного артрита и т.д.) (free radical diseases).
|
|
|
