 |


Особенности строения океанической земной коры
|
|
|
|
Длительное время считалось, что океаническая кора принципиально не отличается по строению от континентальной: океаны (кроме Тихого) представляют собой временно опущенные по разломам блоки, где идет накопление морских осадков, после чего они вновь могут подняться и стать континентами; континентальные же блоки могут опуститься и на них начнется морское
Новые методы изучения дна океанов (глубоководное бурение, геофизические исследования, в том числе использование лазеров) позволили выявить многие детали строения, отличающие океаническую кору от континентальной.
Кора океанического типа состоит из трех слоев (сверху вниз):
Первый слой - осадочная толща мощностью от 0 до 0,5-1 км (в среднем 0,2-0,5 км). Скорость осадконакопления 1-5 мм/тыс, лет. На большей части глубоководного ложа имеет меловой и кайнозойский возраст.
Второй слой - лавы, в том числе подушечные, дайки базальтов (в нижней части слоя).. Мощность 1,5-2 км. Возраст пород средняя юра - кайнозой.
Третий слой - бурением не вскрыт, но драгированием в зонах разломов получены образцы основного (габброиды) и отчасти ультраосновного составов. Средняя мощность 3-4 км-. Третий слой подстилается верхней мантией, сложенной перидотитами. Таким образом, второй и третий слои принадлежат "базальтовому" геофизическому слою.
В пределах Мирового океана выделяются три типа областей: материковые, океанические окраины и ложе океана (Цейслер, 1979).
Материковые окраины. Граница между континентальной и океанической корой не совпадает с географической границей суши и моря. Геологи проводят такую границу в месте сопряжения континентальных и океанических блоков. Периферическим частям материков ниже уровня океана соответствуют шельф и материковый склон. Области шельфа - окраины материков, залитые неглубоким морем (до 250-300 м), сложены корой континентального типа. Ширина шельфа меняется от десятков до более чем 1500 километров (Северный Ледовитый океан). На шельфах продолжает накапливаться осадочный чехол. Поверхность земной коры в области шельфа полого (1-1,5") наклонена в сторону океана. На глубинах 100-200 м обычно располагается бровка шельфа, ниже которой начинается материковый склон. Поверхность последнего наклонена круче (более 3°) и имеет общее ступенчатое строение.
|
|
|
Материковый или континентальный склон - гигантская флексурообразная структура, осложненная системой глобальных разломов. К подножию материкового склона через систему каньонов, часто продолжающих под водой русла крупных рек, мутьевыми потоками и гигантскими оползнями сносятся огромные массы осадков, образующие крупные конусы выноса. Поверхность дна подножия материкового склона обычно обладает чертами аккумулятивной равнины с крупными холмами, которые образованы отдельными конусами выноса или гигантскими оползневыми массивами осадочных пород. Граница континентальной и океанической коры проводится между материковым или континентальным склоном и континентальным подножием.
Океанические окраины. Структурные формы океанических окраин наиболее полно представлены в Тихом океане, где они включают глубоководные окраинные котловинные моря, островные дуги и глубоководные желоба.
Окраинным котловинным морям (Японское, Охотское, Берингово и др.) соответствуют крупные брахиформные депрессии глубиной до 3-5 км. Земная кора окраинных морей местами подобна океанической, однако она нередко обладает увеличенной мощностью осадочного слоя. Поверхность дна окраинных котловинных морей имеет черты аккумулятивной равнины, однако там, где осадков мало, дно обнаруживает высокую степень раздробленности. Строение дна некоторых данных морей осложнено подводными поднятиями (поднятие Ширшова в Беринговоморской котловине, Ямато - в Япономорской и др.), которые возвышаются над дном котловин на 1,5-3 км и имеют блоковое строение.
|
|
|
Островные дуги (Алеутские, Курильские, Японские острова и т.п.) образуют протяженные (1000-3000 км) горные сооружения, которые вместе с сопряженными с ними глубоководными желобами отделяют окраинные котловинные моря от области океанического ложа. Среди островных дуг выделяются два типа: одинарные и двойные. Одинарные дуги образованы узкой (50-70 км) цепочкой вулканических сооружений, покоящихся на низком и широком (до 200 км) сводовом поднятии (Идзу-Бонинская, Марианская и др.), или же широким (70-120 км) и высоким (2-3 км) хребтом, увенчанным цепочкой вулканических аппаратов, смещенных к приматериковому склону дуги (большая часть Алеутской, среднее звено Курило-Камчатской дуги и др.). Вулканизм преобладает базальтовый и андезитовый. Двойные дуги образованы двумя грядами хребтов с одним цоколем. На внешнем хребте вулканизм почти не проявлен; склоны внешнего хребта имеют ступенчато-сбросовое строение.
Островные дуги рассматриваются в качестве геоантиклинальных поднятий современных геосинклинальных областей. Двойные дуги обладают "зрелой" корой материкового типа; в одинарных дугах гранито-гнейсовый слой маломощный.
Глубоководные желоба расположены у подножий мегантиклинориев кайнозойских складчатых систем или на внешних окраинах островных дуг со стороны океана. Глубоководные желоба представляют собой протяженные (1500-4000 км) депрессии глубиной 5-10 км и шириной по дну 5-20 км. Поперечный профиль желобов асимметричный, склоны неравновеликие, ступенчатые, средняя крутизна склонов 5°. Обычно склоны желобов лишены осадочного покрова; последний развит в наиболее глубокой части желобов, куда осадки выносятся мутьевыми потоками. Днища желобов с осадочным выполнением уплощенные.
Глубоководные желоба образуют протяженные системы и главным образом опоясывают центральную часть Тихого океана. Со стороны океанических котловин с желобами граничит такая же протяженная система невысоких пологих краевых валов, являющихся структурными элементами океанского ложа. К склонам желобов и прилегающим островным дугам приурочены зоны повышенной сейсмической активности, которые объясняются выходом на поверхность гигантских поверхностей скола, с которыми связаны как мелко-, так и глубокофокусные землетрясения (зоны Заварицкого-Беньофа). Над глубоководными желобами отмечаются зоны пониженных значений теплового поля.
|
|
|
В представлениях "новой глобальной тектоники" глубоководным желобам отводится важная роль, так как по ним проводятся границы литосферных плит разного типа. Местоположение желоба фиксирует линию'поддвигания океанической плиты под материковую (процесс субдукции).
Ложе океана. Внутриокеанические структурные формы резко различаются степенью подвижности. Среди них выделяются сейсмически активные области (океанические подвижные пояса) и асейсмичные области (океанические платформы или талассократоны). Первая категория структур - глобальная система срединно-океанических хребтов, вторая включает океанические
котловины (плиты), а также различные типы внутриокеанических сводовых и глыбовых поднятий и краевых валов.
Срединно-океанические хребты - протяженная (около 20000 км) и широкая (до 1000 км) система горных сооружений, поднимающихся до 2-3 км над дном океана. Это самые крупные горные системы на поверхности земного шара. Особенно хорошо выражены они в Атлантическом и Индийском океанах. Отдельным вершинам хребтов соответствуют острова вулканического происхождения (о-ва Пасхи, Св.Елены, Св.Павла, Тристан-да-Кунья и др.). Нижние части склонов хребтов обычно пологие, в приосевой части склоны изборождены глубокими продольными желобами и возвышающимися над ними гребнями. Вдоль осевой части хребтов располагается система рифтов.
Рифты представляют собой грабенообразные структуры, в которых центральные блоки ограничены глубинными разломами, доходящими обычно до мантии. Осадочный покров на склонах хребтов крайне маломощен, утонен также и "базальтовый слой". Рифтовые системы отличаются высокой тектонической и вулканической активностью, повышенными значениями теплового поля (в 5-7 раз выше среднего). Мантия в этих зонах ближе подходит к поверхности, а мантийные конвекционные потоки под хребтами направлены снизу вверх. К хребтам приурочены системы полосовых магнитных аномалий. По мнению сторонников гипотезы разрастания океанического дна, в зоне срединно-океанических хребтов происходит формирование новой земной коры за счет выплавки базальтов и раздвижения соседних литосферных плит по горизонтали (спрединг).
|
|
|
Рифтовые системы наблюдаются не только в океанах, они прослеживаются и на континентах. Таковы, например, Восточно-Африканская, Калифорнийская, Байкальская и другие рифтовые области.
Отдельные отрезки подводных хребтов ступенчато смещены в плане по поперечным, так называемым трансформным разломам. На поверхности они выражены уступами дна или узкими глубокими каньонами.
Огромные площади за пределами срединно-океанических хребтов заняты океаническими котловинами с глубинами 4-6 км. Их называют еще океаническими платформами, или талассо-кратонами. Поверхность дна этих депрессий слабо холмистая, с отдельными вулканическими конусами. Многочисленные разломы обусловливают ступенчатый рельеф дна многих котловин. Мощность осадочного покрова в океанических котловинах небольшая, менее 1000 м. Возраст осадков различен. Их геофизическая характеристика подобна характеристикам котловин окраинных морей.
Среди асейсмичных поднятий, разделяющих океанические котловины и осложняющих их строение, выделяется несколько типов: сводовые валообразные поднятия (Гавайский вал, вал Шатского и др.) и глыбовые хребты (Восточно-Индийский и др.). Склоны и сводовые части поднятий увенчаны вулканическими конусами.
В западной части Тихого океана особенно многочисленны подводные горы с плоскими вершинами, погруженными, либо выходящими на поверхность в виде островов. Такие подводные горы образуют целые системы и получили название гайоты.
В океанических структурах развиты лавы основного и ультраосновного состава, причем наиболее широко распространены щелочные базальты.
Дальнейшее изучение структурных форм океанов, их сравнение со структурами континентов, выяснение места океанических структур в общем эволюционном ряду структурных форм земной коры является первостепенной задачей, от решения которой зависит разработка современных геотектонических концепций.
Глава 5
ПРОИСХОЖДЕНИЕ ЖИЗНИ. ПЕРВЫЕ ЭТАПЫ РАЗВИТИЯ ОРГАНИЧЕСКОГО МИРА
|
|
|
Вопрос о происхождении жизни относится к главной проблеме, интересующей людей на протяжении последних столетий. Исследователи, стоящие на научных позициях, должны, прежде всего, решить вопрос: возможно ли зарождение жизни на Земле естественным путем, или жизнь за-, несена на Землю из космоса.
Живое вещество состоит из органических молекул, весьма разнообразных и сложных, благодаря способности углерода образовывать крупные полимерные цепочки, кольца, присоединяя атомы других элементов, главные из которых водород, кислород, азот, фосфор. Органические молекулы составляют нуклеиновые кислоты, белки, жиры, углеводы и другие соединения, из которых построены живые организмы. Все они являются звеньями пищевой цепочки, другими словами, каждый организм пригоден для пищи какому-либо другому. Это свидетельство в пользу общности происхождения всех разнообразных проявлений жизни.
По мнению академика А.И.Опарина, важнейшим свойством живых организмов является упорядоченный обмен веществ. Кроме этого, живые существа должны обладать механизмом передачи наследственных признаков и воспроизведения себе подобных.
Особенностью живого вещества является его дискретность, выражающаяся на разных уровнях в виде организма, популяции, вида и т.д. Надорганизменные образования гораздо устойчивее, чем отдельные организмы, хотя количество организмов, их обновление прямым образом влияет на эту устойчивость. Размножение организмов должно обеспечиваться сохранением информации ее передачей из поколения в поколение.
Учитывая весьма небольшой набор элементов, из которых состоят органические..молекулы, можно с уверенностью заключить, что строительного материала в первичной атмосфере и гидросфере было вполне достаточно. Это водяные пары, СО2, СО, H2S, NH3, CH4, карбиды и другие распространенные соединения. В процессе экспериментов по моделированию процессов биогенеза ("биопоэза" по Дж.Берналу) было сделано открытие, заключающееся в том, что хаотические соединения элементов вполне могли привести к синтезу органических молекул, но лишь при условии Воздействия жесткого ультрафиолетового космического излучения в качестве источника энергии.. Это может вызвать недоумение, ведь известно, что ультрафиолетовые лучи губительны для всего живого. Но на стадии, когда живое вещество только создавалось, энергия ультрафиолета была непременным условием этого процесса. Первые настоящие живые организмы вынуждены были спрятаться в воде от жестких ультрафиолетовых лучей, свободно достигавших земной поверхности в условиях бескислородной атмосферы.
Многочисленные эксперименты, моделирующие процесс образования "органических" молекул в результате неорганических процессов в условиях первичной бескислородной атмосферы, показали, что это не только возможно, но осуществимо многими способами. Такие процессы были не единичными и случайными, но весьма типичными в тех условиях, подобно образованию капель дождя, кристаллов солей и т.п. Эксперименты показали также (Руттен, 1973, и др.), что вполне возможным без участия организмов был и последующий процесс - формирование из этих первичных "органических" молекул различных полимеров: полипептидов, нуклеиновых кислот и далее белков, i
Следующий шаг - объединение первичных молекул в более крупные частицы и их обособление - коацерватные капли. Этот процесс был назван коацервацией (Бунгенберг де Ионг). Коацерва-
ция, по-видимому, происходила при вспенивании с участием поляризованных молекул. Коацерват-ные капли обязательно должны были обособиться с помощью мембран, позволяющих сохранять ' отличные от окружающей среды состав и энергетический уровень. Мембраны были полупроница- " емыми, обеспечивая обмен веществ с окружающей средой —метаболизм. Поглощение пищи и дыхание обеспечивали организм необходимыми материалами и энергией. Наконец, для продолжения жизни требуется воспроизведение себе подобных с передачей им наследственной информации. Воспроизведение (редупликация) осуществляется по "инструкции", заложенной в полимерных цепочках нуклеиновых кислот, хотя происходили и мутации, позволившие приобретать новые свойства и приведшие к поразительному разнообразию жизненных форм. По мере приобретения всех этих свойств из коацерватных капель, взвешенных в "первичном бульоне", возникли живые орга-. низмы. После накопления достаточного количества кислорода смог возникнуть процесс дыхания и кроме бескислородных (анаэробных) появились кислородные (аэробные) формы жизни.
Переход от бескислородной к кислородной атмосфере начался около 1,8 млрд. лет назад, но только к концу силура (~420 млн. лет назад) кислорода в атмосфере стало достаточно много, чтобы надежно защищать поверхность Земли от губительного для жизни ультрафиолетового излучения.
Первоначально живые клетки не имели ядра (прокариоты); позднее от них произошли клетки с более сложной организацией, имеющие ядро (эукариоты).
Первые организмы были гетеротрофными, то есть могли использовать для питания только органические вещества. Автотрофные организмы, которые могли потреблять неорганические вещества, сформировались позднее, с появлением такого энергетически выгодного процесса, как -фотосинтез. Среди микроорганизмов выделяются фототрофные, использующие солнечную энер-рию, и хемотрофные, пользующиеся энергией химических реакций.
|
Лишь с появлением настоящих живых организмов начался этап развития жизни, который можно зафиксировать находками фоссилий, то есть с использованием палеонтологического метода. { Искать следы древней жизни надежнее на территории щитов. Породы докембрия обычно глубоко метаморфизованы, что почти не оставляет шансов на успех. Однако на некоторых фрагментах щитов породы удивительным образом избежали сильного метаморфизма. Например, надсерия Свазиленд (возраст 3,2 млрд. лет) на юге Африки, некоторые районы Канадского щита (2,7 млрд. лет) и др. j Древнейшие свидетельства жизни - это молекулярные ископаемые, то есть фрагменты крупных органических молекул, входивших когда-то в состав организма. Конечно, невозможно определить их принадлежность, они отражают сам факт наличия жизни. К таким ископаемым могут, в частности, принадлежать углистые микроструктуры.
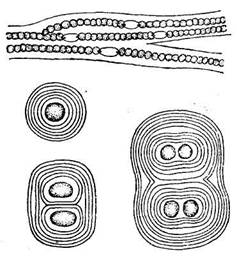
| Рис. 34. Древнейшие прокариоты - цианобионты (сине-зеленые водоросли) - в виде шариков и нитей (увеличено). (По Б.А.Келлеру, 1948) |
| Древнейшими организмами, очевидно, были представители прокариот - бактерии, а также цианобионты, или, как их иногда продолжают называть по старинке, сине-зеленые водоросли, хотя это не водоросли и вообще не растения (рис. 34). С растениями цианобионтов сближает наличие хлорофилла. Именно эти организмы впервые начали выделять кислород в процессе фотосинтеза. Бактерии и цианобионты - важнейшие организмы докембрия, имеющие значение для анализа первых этапов развития органического мира. С их жизнедеятельностью связаны формирование кислородной атмосферы, общий круговорот углерода, накопление карбонатных и железорудных формаций (джеспилитов), пирита, серы, фосфоритов и др. Фоссилизирован-ные остатки бактерий обнаружены в кремнистых породах возраста 3,5 млрд. лет, проблематичные находки имеют воз-
раст Около 3,8 млрд. лет. Примерно такими же возрастными отметками датируются находки циа-нобионтов.
Слизистые нити цианобионтов не могли сохраниться в ископаемом состоянии, однако находят карбонатные пластинки - следы их жизнедеятельности, - которые называются строматолитами. Они бывают пластовой, желваковидной, столбчатой формы. Другая разновидность - отолиты - представлена небольшими сферическими концентрически-скорлуповатыми образованиями, возникшими, к примеру, путем обрастания песчинки или небольшого камешка, тогда как строматолиты сплошь покрывали какую-либо поверхность. Корочки строматолитов, нарастая друг на друга, формировали карбонатные толщи мощностью сотни метров. Кроме этих двух форм встречаются катаграфии - неслоистые узорчатые образования, состоящие из слипшихся комочков.
Первые древнейшие настоящие ископаемые были найдены в формации Ганфлинт на Канадском щите (Онтарио) возраста около 2 млрд. лет. Там были встречены микроостатки разнообразной формы - нитчатые, округлые, с внутренними перегородками. Это свидетельство уже довольно высокой организации жизни.
Остатки микроорганизмов, состоящих из преобразованного фоссилизацией органического кислотоустойчивого вещества, округлой формы, сплющенные либо объемные, размером 3-30 мкм - это мик-рофитофоссилии, которые еще называют акритархами. Они являются, скорее всего, остаткам^ одноклеточных водорослей. Особенно много их в отложениях позднего докембрия, начиная со среднего рифея (1400 млн. лет). Примерно в это же время появились многоклеточные водоросли и грибы. Приблизительно на рубеже среднего и позднего рифея (1 млрд. лет) - первые многоклеточные животные (рис. 35). Возможно, одновременно с ними появились известковые водоросли.

Рис. 35. Общая стратиграфическая шкала докембрия России и развитие органического мира. Границы геохронологических подразделений даны по датировкам региональных подразделений докембрия России (по Б.С.Соколову, 1977, с изменениями авторов)
Есть все основания полагать, что к венду (~670-680 млн. лет назад) жизнь в море существовала в достаточно высокоорганизованных формах и была распространена повсеместно. Находки вендских ископаемых чрезвычайно редки, но не потому, что их было мало, а из-за отсутствия у них скелета или панциря. Ископаемые беспозвоночные вендского возраста впервые были найдены в Южной Австралии, в хр.Флиндерс близ Эдиакары, бассейн Аделаида (рис. 36). Затем подобные
находки были сделаны и на других платформах, в том числе и в России, на берегу Белого моря. Эти удивительные ископаемые были изучены и реконструированы русскими учеными Б.С.Соколовым и М.А.Федонкиным. Большая часть организмов представлена кишечнополостными, а также кольчатыми червями и членистоногими; присутствуют также формы неясного систематического положения, в частности петалонамы и др. Многие из них имеют весьма крупные размеры. Растения, по-видимому, представлены низшими формами - водорослями, фитопланктоном. Даже подобные редкие находки свидетельствуют о существовании значительно развитой, разнообразной вендской биоты, которая была бесскелетной и поэтому плохо сохранилась.
Лишь в начале кембрия многие группы животных приобрели способность выделять твердые раковины, панцири, скелеты и т.д., которые сохранялись в ископаемом состоянии гораздо лучше, чем мягкие ткани животных. Причины этого явления не понятны до конца; возможно, такая способность была связана с временным снижением содержания углекислого газа в атмосфере.

|
Рис. 36. Реконструкция фауны Эдиакары (венд, Южная Австралия),
сделанная М. Глесснером и М. Уэйдом (Glaessner, Wade, 1966)
Кишечнополостные: / - Ediacaria flindersi, 2 - Beltanella gilesi, 3 - Medusinites asteroides, 4 - Mawsonites spriggi, 5 - Cyclomedusa davidi, 6 - C. plana, 7 - Conomedusites lobatus, 8 - Rangea longa, 9 - Arborea arborea, 10 - Pteridinium simplex, кольчатые черви: 11 - Spriggina flounderi, 12 - Dickinsonia costata, 13 -D. elongata, 14 - D. tenuis, членистоногие: 15 - Parvancorina minchami, 16 - Praecambridium sigillum, другие организмы: 17 - Tribrachidium heraldicum, 18 - шарообразные студенистые тела
В свете всего изложенного выше об особенностях развития жизни на Земле становится понятным смысл используемого иногда термина криптозой (греч. криптос - скрытый, тайный; зоэ - жизнь) - "время скрытой жизни", - который был предложен А.Че двиком в 1950 г. как синоним докембрия в противовес его же термину (1930) фанерозой (греч. фанерос - явный, зоэ - жизнь), т.е. "время явной жизни". Фанерозойский эон объединяет палеозойскую, мезозойскую и кайнозойскую эры (греч. палеос - древний, мезос - средний, кайнос - новый, зоэ - жизнь). В последние годы криптозойский эон заменили двумя акронами: архейским или археозойским (греч. архэос древний, зоэ - жизнь) и протерозойским (греч. протерос - первичный, зоэ - жизнь). Видно, что в основу названий всех крупнейших геохронологических подразделений положен единый принцип, основанный на оценке эволюции органического мира.
Глава 6
ДОКЕМБРИЙ. АРХЕЙСКИЙ И ПРОТЕРОЗОЙСКИЙ АКРОНЫ
(АКРОТЕМЫ) - AR + PR
|
|
|
