 |


Семейство белков главного комплекса гистосовместимости
|
|
|
|
Рис. 1-45. Строение иммуноглобулина G.
взаимодействовать с соответствующим (комплементарным) антигеном.
Основные функции антител - обнаружение и связывание чужеродных антигенов, находящихся в организме вне его клеток (в крови, лимфе, межклеточной жидкости, в слизистых секретах). Это происходит с помощью специфических антигенсвязывающих участков разных клонов иммуноглобулинов. Кроме, того, благодаря связыванию антигена с антителом облегчается процесс дальнейшего разрушения чужеродных веществ. Специфичность пути разрушения комплекса антиген-антитело зависит от класса антител.
Классы иммуноглобулинов. Существует 5 классов тяжёлых цепей иммуноглобулинов, отличающихся по строению константных доменов: α, δ, ξ, γ и μ. В соответствии с ними различают 5 классов иммуноглобулинов: A, D, Е, G и М. Особенности строения тяжёлых цепей придают их "шарнирным участкам" и С-концевым областям характерную для каждого класса конформацию. Связывание антигена с антителом изменяет конформацию константных доменов тяжёлых цепей, что определяет путь разрушения комплекса в организме (связывание с белками системы комплемента или поглощение комплекса фагоцитирующими клетками).
Иммуноглобулины М - первый класс антител, синтезирующийся в развивающихся В-лимфоцитах. Различают 2 формы иммуноглобулинов М: мономерная, мембранно-связанная форма и пентамерная, секретируемая В-лимфоцитами в кровь.
Мембранно-с вязанная форма иммуноглобулинов М. Созревающие В-лимфоциты синтезируют мономерные бивалентные молекулы IgM, по структуре похожие на рассматриваемые выше IgG, которые встраиваются в плазматическую мембрану клеток и играют роль первых антиген-распознающих рецепторов. Прикрепление IgM к мембране осуществляется с помощью гидрофобного участка, находящегося в С-концевой ("хвостовой") области тяжёлых цепей, содержащей 25 гидрофобных аминокислотных остатков.
|
|
|
Взаимодействие антигена с рецептором на поверхности В-лимфоцита вызывает его размножение и образование целого клона лимфоцитов, происходящих из одной, стимулированной антигеном клетки. Этот клон В-лимфоцитов будет вырабатывать иммуноглобулины с одинаковыми антигенсвязывающими участками. Однако В-лимфоциты способны переключаться на выработку других классов антител.
Семейство Т-клеточных
антигенраспознающих рецепторов
Если антитела, вырабатываемые В-лимфоцитами, связывают антигены в жидкостях организма (так называемый гуморальный иммунитет), то Т-лимфоциты взаимодействуют с антигенами на поверхности заражённых вирусами и изменённых в результате опухолевой трансформации собственных клеток организма (клеточный иммунитет). Т-лимфоциты узнают антигены только в комплексе с молекулами МНС I или II класса, также присутствующими на клеточной поверхности.
Рецепторы Т-лимфоцитов -гетеродимеры, т.е. состоят из α- и β-цепей. Каждая цепь имеет два
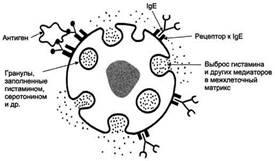
Выброс биологически активных веществ тучной клеткой в результате присоединения антигена к фиксированным на её поверхности IgE.
иммуноглобулиноподобных домена: вариабельный (V) и константный (С) (рис. 1-52). С-концевые участки каждой цепи встроены в плазматическую мембрану. Единственный антигенсвязьшающий участок располагается между двумя вариабельными доменами Vα и Vβ. Количество рецепторов Т-лимфоцитов с разными антигенсвязьшающими участками сопоставимо с разнообразием иммуноглобулинов.
Семейство белков главного комплекса гистосовместимости
Белки главного комплекса гистосовместимости были открыты при изучении вопросов внутривидовой пересадки тканей, откуда и произошло их название. Их называют также белками МНС (см. выше), или белками HLA (от англ. human lymphocyte antigen - человеческие лимфоцитарные антигены), так как впервые они были обнаружены на лимфоцитах человека.
|
|
|
Существует два основных класса молекул МНС: I и II. Молекулы МНС класса I расположены на поверхности практически всех клеток организма человека, а белки МНС класса II только на определённых клетках иммунной системы, называемых антигенпредставляющими клетками. К ним, в первую очередь, относят макрофаги и В-лимфоциты, контактировавшие с антигеном.
Молекулы МНС класса I -гетеродимеры. Они имеют одну полипептидную a-цепь, связанную нековалентными связями с небольшим внеклеточным белком β2-микроглобулином. Полипептидная α-цепь имеет три внеклеточных глобулярных домена (α1, α2, α3), трансмембранный участок и карбоксильный конец, локализованный в цитоплазме (рис. 1-53, А). α3-Домен и β2-микроглобулин имеют конформацию, напоминающую структуру иммуноглобулинов. Домены α1 и α2 содержат вариабельные участки, способные связывать "развёрнутый" антиген (чаще всего пептидный фрагмент чужеродного белка), расположенный на поверхности клеток.
Молекулы МНС класса II -также гетеродимеры. Они состоят из двух полипептидных цепей - α и β, имеющих по одному консервативному иммуноглобулинподобному домену и по одному вариабельному домену на N-концевых участках. Связывание антигенов происходит в области вариабельных доменов α- и β-цепей (рис. 1-53, Б).

Рис. 1-53. Строение белков главного комплекса гистосовместимости: МНС класса I (А) и МНС класса II (Б).
Чужеродные белки в клетке человека (например, белки вирусных частиц), в лизосомах подвергаются ограниченному протеолизу, и небольшие фрагменты этих белков вместе с белками МНС класса I или II экспонируются на поверхности клеточной мембраны.
Комплексы пептид-белок МНС узнаются рецепторами Т-лимфоцитов. В результате происходит специфическое взаимодействие (рис. 1-54), активация Т-лимфоцита и развитие иммунной реакции. Так, взаимодействие цитотоксического Т-лимфоцита с комплексом антиген-МНС I на поверхности заражённой вирусом клетки приводит к высвобождению лимфоцитом специальных

|
|
|
