 |


Макромолекулярная структура РНК.
|
|
|
|
Задание 1. Рассмотрите график плавления ДНК (рис. 2.1.). Объясните зависимость структуры молекулы ДНК от температуры. Объясните, в чем причина данной зависимости. Объясните, почему данный процесс называют кооперативным и обратимым.
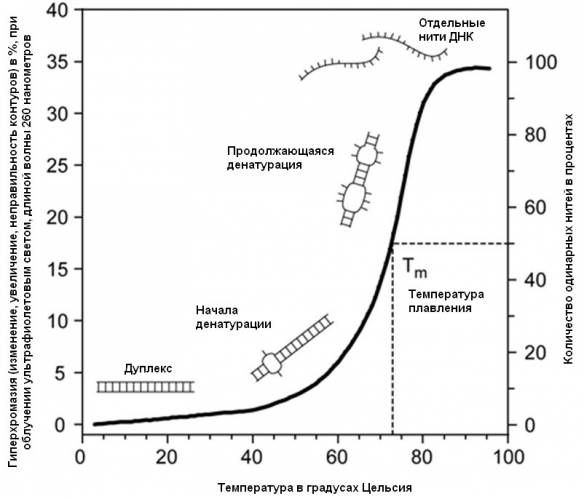
Рисунок 2.1. График плавления ДНК.
Задание 2. Дайте определение следующим терминам: плавление (днатурация) ДНК, «отжиг» (реассоциация, ренатурация) ДНК.
Задание 3. Рассмотрите кривые ренатурации ДНК, выделенной из мыши и ДНК из Е. coli (рис. 2.2.). Фрагменты денатурированной ДНК размером 400 п. н. отжигали при разных величинах C0t и определяли количество ренатурированной ДНК с помощью хроматографии на гидроксилапатите. Объясните, почему ДНК Е. coli денатурирует в узком интервале C0t, а ДНК мыши распадается на несколько кинетических компонентов.
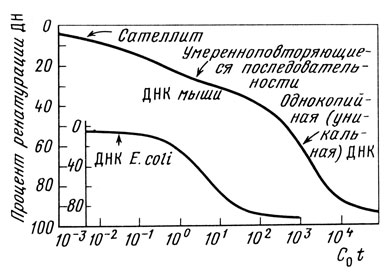
Рисунок 2.2. Кривые ренатурации ДНК, выделенной из мыши и ДНК из Е. coli.
Повторяющиеся последовательности. Одним из фундаментальных отличий организации геномов эукариот является существование в нем многочисленных повторяющихся последовательностей. Эти работы были выполнены группой Р. Браттена (США). Авторы изучали процессы ренатурации и гибридизации нуклеиновых кислот.
Ранее П. Доти (США) и его сотрудники показали, что двойную спираль ДНК можно расплетать или денатурировать, применяя различные воздействия (повышение температуры, изменение рН и др.). В то же время, если инкубировать денатурированную одноцепочечную ДНК при температуре ниже той, которая вызывает денатурацию, комплементарные цепи реагируют между собою, вновь образуя двойную спираль,- происходит ренатурация ДНК. Если к ДНК добавить комплементарную РНК, может происходить образование гибридов ДНК-РНК - гибридизация, которая в дальнейшем стала одним из мощнейших методов изучения нуклеиновых кислот, позволяя выявлять комплементарные молекулы.
|
|
|
Ренатурация нуклеиновых кислот идет в два этапа. На первом в результате случайных столкновений одно-цепочечных молекул происходит правильное соединение двух коротких комплементарных отрезков, а затем, как у молнии, идет быстрое "схлопывание" по всей длине с образованием полной двухцепочечной молекулы. Обычно скорость реакции лимитирует ее первая стадия, или стадия нуклеации. Естественно, что чем выше концентрация ДНК и чем она однороднее, тем чаще будут сталкиваться комплементарные цепи и тем быстрее будет идти реакция. Если весь геном организма уникален и в нем нет одинаковых участков, то скорость ренатурации при прочих равных условиях будет находиться в обратной зависимости от размеров генома. Для безъядерных организмов (прокариот), как показали Бриттен и сотр., это действительно так. Логарифм произведения исходной концентрации ДНК (С0) на время, необходимое для полуренатурации ДНК ч (t1/2), прямо пропорционален размеру генома.
Однако у ядерных организмов, эукариот, как показали те же авторы, это правило нарушается. Содержание ДНК на клетку у этих организмов, размер их генома в десятки и сотни раз больше, чем у бактерий. Однако значительная часть всей ДНК, от 15 до 70 %, ренатурирует гораздо быстрее, чем остальная часть ДНК, со скоростью большей или равной таковой у бактерий. Был сделан вывод, что некоторые отрезки ДНК у эукариот многократно повторяются в геноме.
Дальнейшие исследования позволили разделить геном на три класса последовательностей: сателлитную ДНК, умеренно повторяющиеся последовательности, рассеянные по геному, уникальную ДНК.
Сателлитная ДНК представлена в геноме простыми последовательностями длиной от нескольких до нескольких сот нуклеотидов, повторенными сотни тысяч, а иногда и миллионы раз и сблокированными вместе, т. е. образующими в хромосомах однородные участки. Так, у Drosophila virilis повторяются 7-нуклеотидные сегменты АСАААСТ, АТАААСТ или АСАААТТ, образующие три разных сателлита. Сателлитная ДНК обычно концентрируется в так называемых центромерах и теломерах. Центромеры - это участки, по которым митотические хромосомы прикрепляются к нитям веретена. Теломеры - это концевые участки хромосом. И те и другие, очевидно, не несут генетической информации, но играют структурную роль в организации хромосом и их правильном разделении при митозе. В этих процессах, вероятно, и участвуют простые последовательности сателлитной ДНК.
|
|
|
Умеренно повторяющиеся последовательности составляют от 10 до 50 % всего генетического аппарата у разных видов эукариот. Они рассеяны по геному, образуя отрезки длиной от нескольких сот до нескольких тысяч нуклеотидов. Сильно варьирует и степень их повторяемости. Одни семейства состоят из нескольких десятков, тогда как другие - из сотен тысяч членов. В отличие от сателлитной ДНК, которая инертна, умеренные повторы активно транскрибируются в клетке, т. е. на их матрице идет активный синтез РНК.
Остальная часть генома представлена уникальными последовательностями, т. е. отрезками ДНК, встречающимися в геноме один или небольшое число раз. Скорость ренатурации таких ДНК ниже и соответствует размерам генома.
Три класса ДНК хорошо выявляются на кривых ее ренатурации, если откладывать на оси абсцисс логарифмы C0t, т. е. произведения концентрации ДНК (моль Р/л) на время (с), а по оси ординат - процент ренатурированной ДНК. Например, у млекопитающих около 15 % ДНК ренатурирует в интервале C0t от 10-5 до 10-3 (сателлитная ДНК), еще 10-20 % - в интервале C0t от 10-2 до 101 (умеренные повторы) и остальные ~70 % (уникальная ДНК) - в интервале от 102 до 104.
Существование большого числа повторяющихся нуклеотидных последовательностей в геноме - это вторая универсальная черта в организации генетического аппарата эукариотических организмов, отличающая их от бактерий и вирусов.
В дальнейшем Р. Бриттен и присоединившийся к нему Э. Дэвидсон более подробно изучили свойства умеренных повторов в геноме. Было выявлено два основных класса таких повторов: короткие, длиной в 100-300 пар нуклеотидов (п. н.), и длинные - несколько тысяч пар нуклеотидов (т. п. н.). У большинства видов, например у многих земноводных и млекопитающих, преобладают короткие повторы, перемежающиеся с отрезками уникальной ДНК длиной около 1 т. п. н., тогда как у некоторых других (например, у D. melanogaster) короткие повторы не выявлялись, а вместо них умеренно повторяющаяся ДНК входила в состав длинных отрезков генома (несколько т. п. н.), перемежаясь с участками уникальной ДНК длиной несколько десятков т. п. н. Авторы развили представление о двух основных типах организации генома: подобному геному Xenopus (в основном короткие повторы) и подобному геному дрозофилы (длинные повторы). Природа как коротких, так и длинных повторов была загадочной. Лишь небольшая часть их приходилась на повторяющиеся гены, например, гены для рРНК или для гистонов. Большинство же их как будто не несло никакой генетической информации. Этот парадокс организации эукариотического генома также не был решен до внедрения методов генной инженерии.
|
|
|
По мере накопления общих данных о структуре ДНК и РНК эукариотических клеток начались попытки их обобщения, и появились гипотезы, объясняющие их особенности по сравнению с ДНК и РНК прокариот.
Выше упоминалась уже концепция про-мРНК, согласно которой в ядре происходит синтез высокомолекулярного предшественника мРНК, который затем подвергается частичному разрушению в клеточном ядре, в результате чего образуется зрелая мРНК. Последняя транспортируется из ядра в цитоплазму. Про-мРНК в среднем в 5 раз длиннее зрелой мРНК.
Эта концепция была выдвинута вначале О. П. Самариной, а затем и рядом других авторов. Она стала далее объектом ожесточенных споров, так как появились утверждения, что на самом деле вся дРНК в ядре подвергается распаду и не является предшественником мРНК. Последняя, по мнению соответствующих авторов, синтезировалась сразу, как таковая, и быстро переносилась в цитоплазму.
Дискуссии по этому вопросу заняли около 10 лет. В 1966 г. В. Я. Арион получил первые данные, доказывающие происхождение мРНК цитоплазмы из тяжелой ядерной дРНК. Он проводил гибридизацию меченой ядерной дРНК с ДНК, добавляя в смесь в качестве конкурента мРНК цитоплазмы. Оказалось, что мРНК подавляет связывание тяжелой ядерной дРНК с ДНК примерно на 20 %. В то же время если гибридизацию вели между легкой ядерной дРНК и ДНК, то мРНК цитоплазмы подавляла гибридизацию почти полностью. В обратных опытах, когда вели гибридизацию ДНК с меченой мРНК цитоплазмы в присутствии немеченой ядерной дРНК в качестве конкурента, происходило полное подавление реакции как тяжелой, так и легкой ядерной дРНК (см. рис. 2). Иными словами, в составе тяжелой ядерной дРНК присутствуют как последовательности, переносящиеся затем в цитоплазму, так и последовательности, распадающиеся в ядре. Легкая ядерная РНК обогащается первым типом последовательностей. Однако эти выводы были сделаны на основании гибридизации при низких величинах С0t, когда связываются лишь повторяющиеся последовательности. Тем самым прослеживалась судьба лишь части дРНК. Однозначное доказательство концепции про-мРНК было получено опять-таки лишь тогда, когда были развиты методы генной инженерии.
|
|
|
Вторая концепция касалась организации эукариотических генов, или единиц транскрипции. Гипотезы, предложенные Бриттеном с Дэвидсоном, пытались объяснить, с одной стороны, роль повторов и, с другой - причины образования гигантской про-мРНК. Предполагалось, что повторы лежат перед геном и служат сигналами для регуляции транскрипции. Другое предположение состояло в том, что регуляторные последовательности прочитываются РНК-полимеразой, а в дальнейшем подвергаются распаду. Таким образом, постулировалось, что 5'-конец про-мРНК не несет структурной информации и именно он разрушается в процессе созревания мРНК. Эта концепция в основном оказалась неверной, но она послужила развитию исследований по структуре про-мРНК и мРНК, поскольку ряд ее предсказаний мог быть проверен в опытах.
Лабораторная работа № 3.
|
|
|
