 |


Статические и статокинетические рефлексы (Р.Магнус). Саморегуляторные механизмы поддержания равновесия тела.
|
|
|
|
С участием продолговатого и среднего мозга осуществляются перераспределение тонуса мышц в зависимости от положения тела в пространстве, тонические и установочные, а также статокинетические рефлексы (Рис.6).
Статические рефлексы – направленны на поддержание естественной позы, возникают при изменении положения головы или тела в пространстве.
1.Позные (познотонические) – обеспечивают создание естественной позы.
· рефлекс с вестибулярного аппарата на мышцы разгибатели конечностей- обеспечивает их высокий тонус, сохранение положения позы животного теменем и спиной вверх;
· рефлекс с вестибулярного аппарата на мышцы сгибатели конечностей - возникает при положении животного теменем и спиной вниз и проявляется в повышении тонуса мышц сгибателей конечностей;
· рефлекс с рецепторов мышц шеи на мышцы сгибатели задних конечностей и мышцы разгибатели передних конечностей - проявляется при запрокидывании головы в выпрямлении передних конечностей и сгибании задних;
· рефлекс с рецепторов мышц шеи на мышцы разгибатели задних конечностей и сгибатели передних конечностей - проявляется в сгибании передних и разгибании задних конечностей при наклоне головы и шеи;
· рефлекс с рецепторов мышц шеи на мышцы разгибатели конечности одной стороны и мышцы сгибатели противоположной стороны - проявляется при вращении в разгибании конечностей той стороны тела, в которую поворачивается голова, и в сгибании конечностей противоположной стороны.
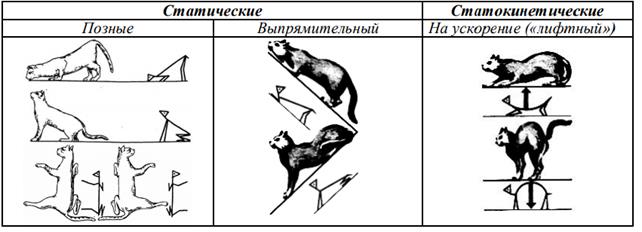
Рис.6. Статические и статокинетические рефлексы.
Выпрямительные рефлексы. Обеспечивают возвращение головы и тела из неестественного положения в естественное. Осуществляются через средний мозг:
|
|
|
· рефлекс с рецепторов вестибулярного аппарата на мышцы головы возникает при положении головы и туловища на боку - проявляется в перераспределении тонуса мышц головы и возврате головы в естественное положение;
· рефлекс с тактильных рецепторов кожи при положении животного лежа на боку на мышцы головы - обеспечивает возврат головы в естественное положение;
· рефлекс с рецепторов мышц шеи, возникающий при изменении положения шеи, на мышцы туловища - обеспечивает перевод туловища в положение, соответствующее положению шеи, за счет перераспределения тонуса мышц;
· рефлекс с рецепторов кожи туловища, возникающий при положении животного на боку, на мышцы туловища- обеспечивает за счет перераспределения тонуса мышц возврат туловища в естественное положение, соответствующее положению головы и шеи.
Статокинетические рефлексы – направлены на поддержание позы и равновесия при перемещении тела в пространстве с ускорением или вращением, когда происходит перераспределение тонуса мышц глаз, туловища и конечностей, что обеспечивает устойчивое положение глаз, головы и тела в пространстве:
· рефлекс с рецепторов мышц одной конечности на мышцы других - возникает при ходьбе животного, когда при сгибании одной конечности повышается тонус мышц разгибателей других трех конечностей;
· рефлекс «нистагм головы» возникает при вращательных движениях - проявляется в движении головы в противоположную сторону вращения туловища, а затем в скачкообразном перемещении головы в положение, соответствующее положению туловища;
· рефлекс «нистагм глаз» возникает при вращательных движениях - проявляется в движении глаз в противоположную сторону вращения головы и туловища, а затем в скачкообразном перемещении глаз в положение, соответствующее положению туловища;
· лифтовый рефлекс возникает при линейном ускорении движения вверх и вниз; в первом случае повышается тонус мышц разгибателей, во втором — тонус мышц сгибателей.
|
|
|
1.9. Физиология мозжечка, его влияние на моторные и вегетативные функции организма.
Мозжечок — одна из интегративных структур головного мозга, принимающая участие в координации и регуляции произвольных и непроизвольных движений, вегетативных и поведенческих функций (Рис.7).
Мозжечок анатомически и функционально делится на архи-, палео-и неоцеребеллюм.
1.Архицеребеллюм (древний мозжечок) — имеет наиболее выраженные связи с вестибулярной системой, что объясняет значение мозжечка в регуляции равновесия.
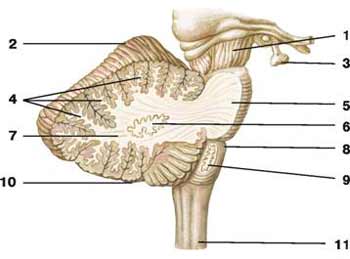
Рис. 7. Мозжечок.
1 - ножка мозга; 2 - верхняя поверхность полушария мозжечка; 3 - гипофиз;
4 - белые пластинки; 5 - мост; 6 - зубчатое ядро; 7 - белое вещество; 8 -продолговатый мозг; 9 -ядро оливы;10 - нижняя поверхность полушария мозжечка; 11 - спинной мозг.
Архицеребеллум получает информацию о пространственном положении головы от вестибулярной системы и о движениях головы посредством кинетических импульсов от рецепторов полукружных каналов. Это позволяет мозжечку синергично модулировать спинномозговые двигательные импульсы, что обеспечивает поддержание равновесия вне зависимости от положения или движений тела. Повреждение клочково-узелковой зоны ведет к нарушению равновесия и неустойчивости при стоянии (астазия) и ходьбе (абазия). Мозжечковая атаксия не увеличивается при закрытых глазах в противоположность атаксии, вызванной поражением задних канатиков спинного мозга. Мозжечковая атаксия является результатом неспособности мышечных групп к координированному действию – асинергия (Рис.8).
Повреждение клочково-узелковой зоны нарушает реакцию на калорическую и ротаторную пробы, применяемые при проверке вестибулярной функции. Аналогичная утрата функции вызывается прерыванием путей, идущих к клочково узелковой зоне или от нее. Равновесие поддерживается следующей рефлекторной дугой: импульсы, начинающиеся в лабиринте, идут как прямо, так и опосредованно через вестибулярные ядра в архицеребеллум и затем к ядрам шатра, откуда эфферентные импульсы возвращаются к латеральным вестибулярным ядрам и далее – в ретикулярную формацию. Через преддверно спинномозговые и ретикулярно спинномозговые пути, задний продольный пучок импульсы достигают клеток передних рогов и модулируют их активность.
|
|
|
2. Палеоцеребеллюм (старый мозжечок) — получает информацию преимущественно от проприорецептивных систем мышц, сухожилий, надкостницы, оболочек суставов.
Палеоцеребеллум получает афферентные импульсы от спинного мозга через передние и задние спинно мозжечковые и от дополнительного клиновидного ядра через клиновидно мозжечковый путь. Эфферентные импульсы от палеоцеребеллума модулируют активность антигравитационной мускулатуры и обеспечивают достаточный для прямостояния и прямохождения мышечный тонус. Спинномозговые импульсы проецируются в кору палеоцеребеллума в соматотопическом порядке, отражая в каждом из полушарий мозжечка ипсилатеральную половину тела. Кора области, прилежащей к червю, «проецируется» в пробковидное и шаровидное ядра, кора червя – в ядро шатра. Поражение палеоцеребеллума вызывает туловищную атаксию. Однако поражение редко ограничивается палеоцеребеллумом, так как имеется некоторое функциональное перекрытие между палеоцеребеллумом и неоцеребеллумом. Поэтому во многих случаях невозможно считать ту или иную клиническую симптоматику проявлением поражения ограниченной области мозжечка.
3. Неоцеребеллюм (новый мозжечок) —получает информацию от коры, преимущественно по лобно-мосто- мозжечковому пути, от зрительных и слуховых репетирующих систем. Это свидетельствует о его участии в анализе зрительных и слуховых сигналов и в организации соответствующих реакций.

Рис.8. Очаговые поражения мозжечка.
Преимущественное поражение полушарий мозжечка (неоцеребеллума) ведёт к расстройству его противоинерционных влияний и, в частности, к возникновению динамической атаксии. Она проявляется неловкостью движений конечностей, которая оказывается особенно выраженной при движениях, требующих точности.
Функции мозжечка. Функции мозжечка формируют три главных его влияния на организм (Л.А.Орбели, 1940): на двигательный аппарат, афферентные системы и вегетативную нервную систему.
|
|
|
1.Двигательные функции мозжечка. Выделяют следующие функции:
Регуляция мышечного тонуса, позы и равновесия осуществляется преимущественно древним мозжечком (флоккулонодулярная доля) и частично старым мозжечком, входящими в медиальную червячную зону. Получая и обрабатывая импульсацию от вестибулярных рецепторов (по вестибуломозжечковому пути), от проприорецепторов аппарата движения и рецепторов кожи (по спинно-мозжечковым путям), от зрительных и слуховых рецепторов (через четверохолмие по тектоцеребеллярным путям), мозжечок способен оценить состояние мышц, положение тела в пространстве и через ядра шатра, используя вестибуло-, ретикуло- и руброспинальный тракты, произвести перераспределение мышечного тонуса, изменить позу тела и сохранить равновесие.
Координация позы и выполняемого целенаправленного движения осуществляется старым и новым мозжечком, входящими в про межуточную (околочервячную) зону. В кору этой части мозжечка поступает импульсация от рецепторов аппарата движения, а также импульсация от моторной коры (программа произвольного движения).
Участие в программировании целенаправленных движений осуществляется новым мозжечком — той частью полушарий мозжечка, которая входит в латеральную зону. Кора этой части мозжечка получает импульсацию преимущественно из ассоциативных зон коры большого мозга через ядра моста. Эта информация характеризует замысел движения.
Наряду с этим полушария мозжечка осуществляют функцию инициации движения.
2.Афферентная функция мозжечка. В реализации влияния мозжечка на афферентные системы организма, без сомнения, большую роль играют проекции ядер мозжечка на специфические и неспецифические ядра таламуса как главного центра переключения в сенсорных системах. Можно предположить, что механизмы влияния мозжечка на сенсорные функции связаны с его влиянием на эфферентный контроль активности рецепторного аппарата и центров переключения в сенсорных системах.
3.Регуляция вегетативных функций. Л.А.Орбели установил, что мозжечок влияет и на различные вегетативные функции. Эти влияния могут быть возбуждающими и тормозящими. Например, при раздражении мозжечка увеличивается или снижается кровяное давление, изменяется частота сердцебиений, дыхание, пищеварение. Мозжечок влияет на обмен веществ. На эти функции он воздействует через вегетативные нервные центры, координируя их активность с движением. Функции внутренних органов изменяются в связи с изменением обменных процессов в них. Поэтому мозжечок оказывает на них адаптационно-трофическое влияние.
|
|
|
1.10. Особенности нейронной организации ретикулярной формации ствола мозга и ее нисходящее влияние на рефлекторную деятельность спинного мозга. Восходящие активирующие влияния ретикулярной формации ствола мозга на кору больших полушарий. Участие ретикулярной формации в формировании целостной деятельности организма.
Ретикулярная формация (РФ) образована совокупностью нейронов, расположенных в его центральных отделах как диффузно, так и в виде ядер. Ретикулярная формация объединяет различные участки ствола мозга (ретикулярную формацию продолговатого мозга, варолиева моста и среднего мозга). В функциональном отношении в ретикулярной формации разных отделов мозга есть много общего, поэтому целесообразно рассматривать ее как единую структуру. Ретикулярная формация представляет собой диффузное накопление клеток разного вида и величины, которые разделены многими волокнами. Кроме этого, в середине ретикулярной формации выделяют около 40 ядер и подьядер. Нейроны ретикулярной формации имеют широко разветвленные дендриты и продолговатые аксоны, часть которых делится Т-образно (один отросток направлен вниз, образуя ретикулярный-спинальный путь, а второй - в верхние отделы головного мозга).
Свойства ретикулярных нейронов. Нейроны ретикулярной формации имеют длинные маловетвящиеся дендриты и хорошо ветвящиеся аксоны, кото рые часто образуют Т-образное ветвление: одна из ветвей аксона имеет нисходящее, а вторая — восходящее направления (Рис.9). Ветви нейронов под микроскопом образуют сеточку (ретикулум), с чем и связано название данной структуры мозга, предложенное О. Дейтерсом (1865).
Рис. 9.Нейронная организация ретикулярной формации.
А. Нейрон ретикулярной формации.
Б. Коллатерали (по М. Э. Шейбэлу и А. Б.Шейбэлу)
Функциональные особенности ретикулярных нейронов:
· полисенсорная конвергенция: принимают коллатерали от нескольких сенсорных путей, идущих от разных рецепторов. В основном это полимодальные нейроны, имеющие большие рецепторные поля.
· у нейронов РФ длительный латентный период ответа на периферическую стиму ляцию в связи с проведением возбуждения к ним через многочисленные синапсы.
· они имеют тоническую активность, в покое 5—10 имп/с.
· нейроны РФ обладают высокой чувстви тельностью к некоторым веществам крови (например, к адреналину, СО2) и лекарст вам (к барбитуратам, аминазину и др.).
Ретикулярная формация имеет связи с многочисленными структурами ЦНС. Афферентные входы поступают в латеральные области РФ преимущественно от трех источников:
• температурных и болевых рецепторов по волокнам спиноретикулярного тракта и тройничного нерва импульсация поступает в ретикулярные ядра продолговатого мозга и моста;
• сенсорной и, частично, других зон коры головного мозга по кортикоретикулярным путям импульсация поступает в ядра, дающие начало ретикулоспинальным трактам (гигантоклеточное ядро, оральное и каудальное ядра моста), а также в ядра, которые проецируются на мозжечок (парамедианное ядро и ядро покрышки моста);
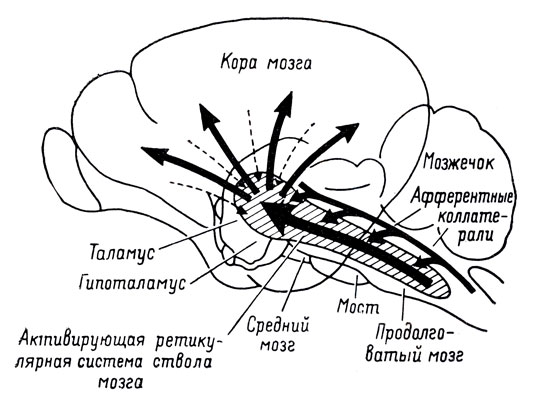
Рис. 10. Схематическое изображение мозга кошки, показывающее расположение ретикулярной формации, распределение коллатералей афферентных путей и активирующие кортикальные проекции (по Т. Э.Старцлу, С. У. Тэйлору и Г. У.Мэгуну).
• ядер мозжечка по мозжечково-ретикулярному пути импульсация поступает в гигантоклеточное и парамедианное ядра и ядра моста. Эфферентные выходы формируются преимущественно в медиальных областях РФ и проецируются:
• к спинному мозгу по латеральному ретикулоспинальному тракту (от гигантоклеточного ядра) и по медиальному ретикулоспинальному тракту (от каудального и орального ядер моста);
• к верхним отделам головного мозга (не специфическим ядрам таламуса, заднему гипоталамусу, полосатому телу) идут восходящие пути, начинающиеся в ядрах продолговатого мозга (гигантоклеточном, латеральном и вентральном) и в ядрах моста;
• к мозжечку идут пути, которые начинаются в латеральном и парамедианном ретикулярных ядрах и в ядре покрышки моста.
Нейроны и ядра ретикулярной формации (РФ) входят в состав центров, регулирующих функции внутренних органов (кровообращения, дыхания, пищеварения), тонус скелетной мускулатуры, активность коры большого мозга. Обширны связи ретикулярной формации с другими отделами ЦНС и рефлексогенными зонами: она получает импульсацию от различных рецепторов организма и отделов ЦНС и в свою очередь посылает импульсы во все отделы мозга. При этом выделяют восходящие и нисходящие влияния ретикулярной формации (Рис.10).
Можно выделить следующие влияния ретикулярной формации:
1. Специфические (ядра продолговатого мозга).
2. Неспецифические:
· восходящие влияния РФ на большой мозг могут быть как активирующими, так и тормозными. Импульсы ретикулярных нейронов продолговатого мозга, моста и среднего мозга поступают к неспецифическим ядрам таламуса и после переключения в них проецируются в различные области коры. Кроме таламуса восходящие влияния поступают также в задний гипоталамус, полосатое тело.
· нисходящие влияния ретикулярной формации на моторные спинальные центры. От ретикулярного гигантоклеточного ядра продолговатого мозга идет частично перекрещенный латеральный ретикулоспинальный тракт, волокна которого оканчиваются на вставочных нейронах спинного мозга. Через эти интернейроны они возбуждают α- и γ-мотонейроны мышц-сгибателей мускулатуры конечностей и реципрокно тормозят с помощью тормозных интернейронов мышцы-разгибатели.
Одним из основных проявлений повреждения ретикулярных структур у человека является потеря сознания. Она бывает при черепно-мозговых травмах, нарушении мозгового кровообращения, опухолях и инфекционных процессах в стволе мозга. Длительность состояния обморока зависит от характера и выраженности нарушений функции ретикулярной активизирующей системы и колеблется от нескольких секунд до многих месяцев. Дисфункция восходящих ретикулярных влияний проявляется тоже потерей бодрости, постоянной патологической сонливостью или частыми приступами засыпания (пароксизмальная гиперсомия), беспокойным ночным сном. Наблюдаются также нарушения (чаще повышении) мышечного тонуса, различные вегетативные изменения, эмоционально-психические расстройства и др.
1.11. Гипоталамус. Характеристика основных ядерных групп. Роль гипоталамуса в интеграции вегетативных, соматических и эндокринных функций, в формировании эмоций, мотиваций, стресса, биоритмов.
Гипоталамус — структура промежуточного мозга, организующая эмоциональные, поведенческие, гомеостатические реакции организма.
Функционально ядра гипоталамуса делят на переднюю, среднюю и заднюю группы ядер. Окончательно созревает гипоталамус к 13-14 годам, когда заканчивается формирование гипоталамо-гипофизарных нейросекреторных связей. Мощные афферентные связи гипоталамуса с обонятельным мозгом, базальными ганглиями, таламусом, гиппокампом, орбитальной, височной и теменной корой определяют его информативность о состоянии практически всех структур мозга. В то же время гипоталамус посылает информацию к таламусу, ретикулярной формации, вегетативным центрам ствола и спинного мозга.
Нейроны гипоталамуса имеют особенности, которые определяют специфику функций самого гипоталамуса. К этим особенностям относятся: чувствительность нейронов к составу омывающей их крови, отсутствие гематоэнцефалического барьера между нейронами и кровью, способность нейронов к нейросекреиии пептидов, нейромедиаторов и др. Ядра гипоталамуса взаимосвязаны друг с другом и с другими структурами нервной системы сложной системой афферентных и эфферентных волокон (Рис.11.).

Рис. 11. Гипоталамус и его главные ядра.
1 - передняя спайка; 2 - гипоталамическая борозда; 3 - паравентрикулярное ядро; 4 - дорсомедиальное (верхнемедиальное) гипоталамическое ядро;
5 - группа ядер заднего гипоталамуса; 6- ядра серого бугра; 7 - ядро воронки;
8 - глубление воронки; 9 –воронка; 10 – гипофиз; 11 - перекрест зрительных нервов; 12 - оптическое ядро; 13 - переднее гипоталамическое ядро;
14 - терминальная пластинка.
Функции гипоталамуса. В целом, за счет большого количества входных и выходных связей, полифункциональности структур, гипоталамус выполняет интегрирующую функциювегетативной, соматической и эндокринной регуляции, что проявляется и в организации его ядрами ряда конкретных функций.
1. Вегетативная регуляция. Влияние на симпатическую и парасимпатическую регуляцию позволяет гипоталамусу воздействовать на вегетативные функции организма гуморальным и нервным путями. Возбуждение ядер передней группыгипоталамуса приводит к реакции организма, его систем по парасимпатическому типу, т.е. реакциям, направленным на восстановление и сохранение резервов организма. Возбуждение ядер задней группывызывает симпатические эффекты в работе органов. Стимуляция ядер средней группыгипоталамуса приводит к снижению влияний симпатической системы.
2. Эндокринные функции. Нейроны передней группы ядер гипоталамуса продуцируют так называемые рилизинг-факторы (либерины) и ингибируюшие факторы (статины), которые регулируют активность передней доли гипофиза - аденогипофиз.
3. Интегративная функция. Доказано, что гипоталамус координирует практически все системы органов и управляет сенсорными, двигательными и вегетативными функциями организма.
· Терморегуляция. Передние ядра гипоталамуса регулируют процесс теплоотдачи. При их повреждении теряется способность к адаптации к повышению температуры. Задняя группа ядер контролирует процесс теплообразования. При их повреждении утрачиваются адаптивные реакции на понижение температуры: отсутствует мышечная дрожь, сужение сосудов. Центр терморегуляции возбуждается при изменении температуры окружающей среды за счет возбуждения холодовых и тепловых рецепторов кожи. В гипоталамусе есть отдельные нейроны обладающие термочувствительностью. Они возбуждаются при изменении температуры омывающей их крови.
· Регуляция обменных процессов. При раздражении задних ядер ускоряются обменные процессы в различных органах и тканях. В результате повышается уровень глюкозы в крови (гипергликемия), т. к. стимулируется гидролиз гликогена в печени и мышцах. Также увеличивается сила и частота сердечных сокращений, повышается тонус гладких мышц сосудов, кровяное давление, частота и глубина дыхания.
· Регуляция половой деятельности. Передняя группа ядер гипоталамуса стимулирует процессы полового созревания за счет стимуляции выделения рилизинг-факторов для гонадотропных гормонов передней доли гипофиза. В результате повышается выброс гонадотропных гормонов из аденогипофиза, которые стимулируют развитие ткани гонад и выделение половых гормонов и у женщин и у мужчин. Задняя доля несколько тормозит половые функции. При поражении задней доли (серого бугра) наблюдается ускорение полового созревания.
· Гипоталамус регулирует эмоциональные реакции – в эксперименте у животных вживлялись электроды в различные ядра гипоталамуса: передняя группа ядер - центр положительных эмоций, задняя группа - центр агрессии и отрицательных эмоций.
· Регуляция деятельности системы пищеварения. В гипоталамусе располагаются образования пищевого центра, которые обеспечивают состояние голода и сытости. Латеральные ядра - центр голода, при их раздражении возникает гиперфагия и булемия; при их разрушении - нет чувства голода - смерть от истощения. Вентромедиальные ядра - центр насыщения при их раздражении по принципу реципрокности тормозятся латеральные ядра, возникает состояние сытости. В дорсомедиальных ядрах - центр жажды.
|
|
|
