Цель занятия: изучить основные закономерности модификационной изменчивости организма, овладеть некоторыми методами их статистического анализа.
Предлагается оценить вариабельность нижеследующих антропометрических показателей в группе. Для этого необходимо провести замеры и предварительный расчет индексов каждого студента.
Кормический индекс.
Для определения индекса измеряемый садится на горизонтальную поверхность и измеряется от макушки до плоскости сиденья. Измерение проводится при помощи антропометра, спина должна быть максимально выпрямлена. Далее определяется рост стоя. Кормический индекс рассчитывается по формуле:
(рост сидя / общий рост)х 100%
Определение длины руки.
Общая длина руки измеряется от латеральной точки акромиального отростка лопатки до кончика третьего пальца. При измерении рука с ладонью, повернутой внутрь, вытягивается вдоль тела, чтобы локоть, кисть и пальцы были максимально разогнуты в суставах
Антропометрия головы.
Шведский анатом Ретциус более 100 лет назад предложил рассчитывать головной индекс как показатель формы головы. Калипером измеряется максимальная длина от глабеллы (костного вздутия между надбровными дугами) до описток-раниума (наиболее удаленной точки затылка). Максимальная ширина, измеряемая под прямым углом к сагитальной плоскости, обычно определяется в области теменных бугров. Головной индекс рассчитывается по формуле
(ширина / длина) х 100%
Весь диапазон изменений головного индекса делят на три группы: долихоцефалы (ниже 75%), мезоцефалы (75-80 %) и брахицефалы (выше 80%). Однако головы с одинаковым индексом могут существенно отличаться по своей форме в целом.
С помощью ростомера измерить рост студентов данной группы.
Определить массу тела тех же студентов.
Провести измерения по двум из пяти предложенных антропометрических показателей. Все данные записать в таблицы
Распределить полученные результаты по классам с интервалом 3,0 см. Вычислить среднюю арифметическую (х, у) каждого вариационного ряда, квадрат отклонений от средней арифметической (х. - х) 2 , среднее квадратичное отклонение (б — сигма), коэффициент вариации (V), коэффициент корреляции (г) и ошибку средней арифметической (т).
Среднее квадратичное отклонение дает возможность при сравнении данного признака с другими в значительно большей степени судить о размахе изменчивости (т.е. о норме реакции), при сравнении изменчивости различных признаков (рост, масса тела и др.) необходимо вычислить и коэффициент вариации
V = (б х 100%) / X
Чем больше проведено измерений, тем точнее результат. Поэтому обязательно следует учитывать среднюю ошибку (т). На ошибку влияет и степень изменчивости признака, определяемая средним квадратичным отклонением Дополнительная литература
ИДЕНТИФИКАЦИЯ ГЕНОТИПА ПО ФЕНОТИПУ С ПОМОЩЬЮ КОРРЕЛЯЦИЙ ПРИЗНАКОВ
Г.М.Скуридин, н.с., С.Ф.Коваль, к.б.н., с.н.с.
Институт цитологии и генетики
Информационный вестник ВОГи А №19 2002 год
Модуль 8. Генетика иммунитета
Комплексная цель:
Ознакомиться с историей изучения генетических основ иммунитета. Изучить генетическую основу систем групп крови АВО и резус фактора. Освоить систему HLA. Рассмотреть генетически обусловленную устойчивость к ВИЧ инфекции. Ознакомиться с генетическими аспектами аутоиммунных заболеваний. Изучить генетику антител и системы антиген/рецептор.
Глава 18. Важность клеточной поверхности
Автор: Шкурат Татьяна Павловна
Группы крови системы АВО
Резус-фактор
Комплекс гистосовместимости HLA
Группы крови системы АВО
Из истории. Первые документированные переливания крови проводились ещё в XVII веке, но представляли собой скорее медицинские казусы. Например, французский врач того времени Жан-Батист Дени переливал кровь ягнят и телят буйным умалишенным в надежде, что она своей "мягкостью и свежестью успокоит сердце и кипение крови" больных. Этот метод был запрещён решением французского суда после того, как в результате очередной подобной процедуры один из пациентов умер.
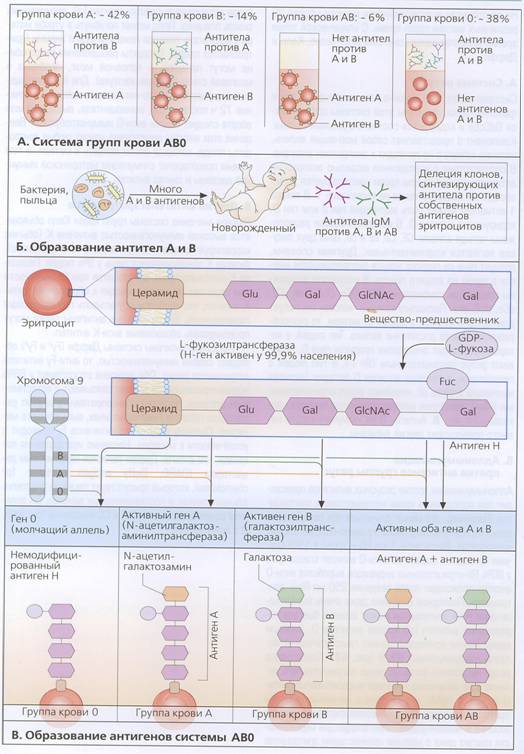
Переливания крови человеку от человека появились на регулярной основе в начале XIX века — в Англии. Остались воспоминания одной из первых пациенток, потерявшей много крови при родах и получившей затем четверть литра донорской крови. По ее словам, она ощутила, "будто сама жизнь проникает в ее организм". Со временем, было установлено, что и переливание крови от одного человека другому вовсе не всегда проходит успешно. Необходимо, чтобы кровь донора "прижилась" в организме того, кому эту кровь вливают (реципиента), оказалась с ней совместимой. Условия подобной совместимости были открыты лишь в начале ХХ века венским исследователем Карлом Ландштейнером. В 1900 году он опубликовал статью, в которой впервые сообщалось об индивидуальных отличиях крови людей. Смешивая взятую у разных лиц сыворотку (жидкую часть) крови со взятыми у других людей эритроцитами (красными клетками крови), он обнаружил, что такое смешение в одних случаях приводило к слипанию эритроцитов и превращению их в сгустки, тогда как в других это явление отсутствовало. На основании подобных экспериментов было установлено, что у людей существуют четыре разных типа крови — четыре группы: 0(I), А(II), В(III) и АВ(IV).
Первая разумная генетическая теория была предложена фон Дунгерном и Гиршфельдом (1911). Для объяснения четырех фенотипов А, В, 0 и АВ они предположили наличие двух независимых пар генов (А,0; В,0), где А и В – доминантные аллели. Бернштейн (1925) проверил эту гипотезу, используя соотношения, ожидаемые из закона Харди-Вайнберга. Он пришел к выводу об ошибочности их концепции и предложил правильное объяснение – три аллеля с шестью генотипами и четырьмя фенотипами вследствие доминирования аллелей А и В над аллелем 0.
Таблица 18.1
Система групп крови АВО
генотип
антиген
фенотип
IA IA
А
А
А
IA I0
IB IB
В
В
В
IB I0
IА IВ
А,В
АВ-
I0 I0
нет
На сегодняшний день известно, что система групп крови АВ0 основана на существовании естественных антител (изоагглютининов) против антигенов групп крови А и В, экспрессирующихся на эритроцитах. Антитела вырабатываются только против антигенов, не экспрессируемых на эритроцитах хозяина. Люди, относящиеся к типу А (около 42% жителей центральной Европы, 41% белого населения Америки, а также 27% афроамериканцев и мексиканцев), имеют антитела против антигена группы крови В. Эти антитела способствуют агглютинации и лизису эритроцитов, несущих антигены В; их называют анти-В-антителами. Если такие эритроциты перелить индивиду с группой крови А, то они немедленно подвергнуться лизису. Люди, относящиеся к типу В (около 14: европейцев, 20 афроамериканцев, 13% мексиканцев, а также 25% азиатов), имеют природные анти-А-антитела. Существуют люди типа АВ (около 6% всего населения), на эритроцитах которых экспрессируются как А, так и В антигены, а в сыворотке нет ни анти-А, ни анти-В антител. У людей с группой крови 0 (около 38% европейцев, 45% белого населения Америки, 49% афроамериканцев, 56% мексиканцев), на эритроцитах нет ни А, ни В антигенов, а в сыворотке присутствуют как анти–А, так и анти-В антитела. Антигены А подразделяются на подтипы А1 (80%) и А2 (29%). Анти А-1 антитела изредка встречаются у людей с типом А2. Антигены экспрессируются главным образом на эритроцитах, но слабая экспрессия наблюдается также на тромбоцитах и эндотелиальных клетках.
Гены АВ и 0 расположены на длинном плече хромосомы 9. Они кодируют гликозилтрансферазы, переносящие различные группы сахаров к веществам-предшественникам. Перенос L-фукозы к цепи предшественника осуществляется продуктом так называемого Н-гена у 99, 9% людей. Это приводит к образованию антигена Н, экспрессирующегося на эритроцитах и состоящего из остатков глюкозы, галактозы, N-ацетилглюкозамина, фукозы. Активный ген А (у людей с типом крови А) кодирует специфическую N-ацетилглюкозаминилтрансферазу, присоединяющую к антигену Н молекулу N-ацетилгалактозамина с образованием антигена А. Если активен ген В, то вместо этого синтезируется молекула галактозилтрансферазы. Иными словами, у людей с группой крови В к антигену Н присоединяется молекула галактозы. У людей с активными генами А и В на эритроцитах происходит экспрессия обоих антигенов. У людей с активным молчащим геном 0 антиген Н остается немодифицированным, что соответствует группе крови 0.
Гены АВ и 0 наследуются в соответствии с законами Менделя. Следовательно, люди типа А могут иметь генотип АА или А0, а люди типа В – ВВ или В0. Индивиды типа АВ имеют генотип АВ, а индивиды типа 0 – генотип 00. Естественные анти-А и анти-В антитела представляют собой главным образом иммуноглобулины класса IgM. Однако, у индивидов с группой крови 0 против А и В антигенов вырабатываются также антитела класса IgG.
Рис. 18.1. Система групп крови АВ0.
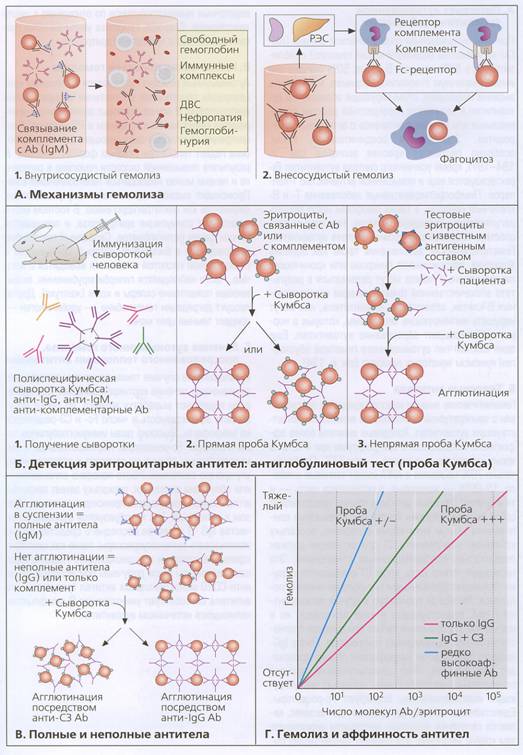
Поскольку анти-А и анти-В изоагглютинины присутствуют в организме от природы, то однократное переливание АВ0-несовместимых эритроцитов может привести к тяжелой форме гемолиза. После первичной встречи с антигенами кроме природных антител может начаться выработка новых иммуноглобулинов.
Фенотип крови Бомбей
Группа крови АВ0 хорошо исследована биохимически. Известно, что антигены А и В представляют собой карбогидратные группы (углеводные остатки), связанные с жирными кислотами, молекулы которых выступают над поверхностью мембраны эритроцитов. Специфичность этих антигенов обусловлена концевым остатком карбогидратной цепи. Оба антигена происходят из одного и того же предшественника, или Н-вещества, к молекулам которого затем добавляется один или два концевых углеводных остатка сахара.
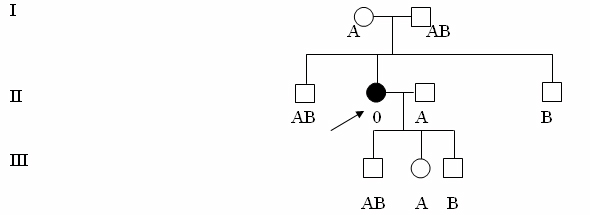
В крайне редких случаях, например, у женщин из Бомбея (Индия) имеется дефектное Н-вещество, которое не связывается с ферментом, добавляющим концевые углеводные остатки. У носителей группы крови 0 это приводит к фенотипу Бомбей. Оказалось, что этот дефект обусловлен рецессивной мутацией h в локусе, не гомологичном локусам А и В. Поэтому при наличии у человека аллелей IA и (или) IB вместе с генотипом hh антигены А и В в крови не обнаруживаются. Вот почему у одной из жительниц Бомбея с генотипом hh определили группу крови 0, хотя ее родители имели группу крови АВ, а сын был носителем аллеля IB.
Рис. 18.2. Часть родословной женщины с фенотипом крови Бомбей. Генетически она имеет группу крови В, но из-за отсутствия антигенов АВ ее кровь отнесена к группе 0.
Опыты Ландштейнера и сегодня используют для определения группы крови пациента. Если, например, не зная групповой принадлежности крови, взять наугад сыворотку группы 0 и эритроциты группы 0 и смешать их, то ничего не произойдёт, поскольку антителам сыворотки не за что ухватиться на эритроцитах 0. Но если ту же сыворотку группы 0 смешать с эритроцитами группы А, то антитела анти-А, присутствующие в сыворотке 0, "схватятся" за молекулы А на эритроцитах группы А и вызовут слипание эритроцитов, собрав их в сгустки. То же самое произойдет и в кровеносных сосудах пациента с группой 0, а в дополнение к этому сработает механизм разрушения эритроцитов, покрытых антителами.
Резус-фактор
История вопроса. В 1939г. Левин и Стетсон исследовали сыворотку крови женщины, которая родила мертвый плод и в анамнезе которой имело место переливание крови мужа, совместимой по АВ0 группе. При этом ими были обнаружены особые антитела. Позже Левин и Стетсон показали, что из 1010 образцов крови только 21 дал отрицательную реакцию с этими антителами. Выявленные антитела никакой связи с системами групп крови АВ0, MN и Р не имели.
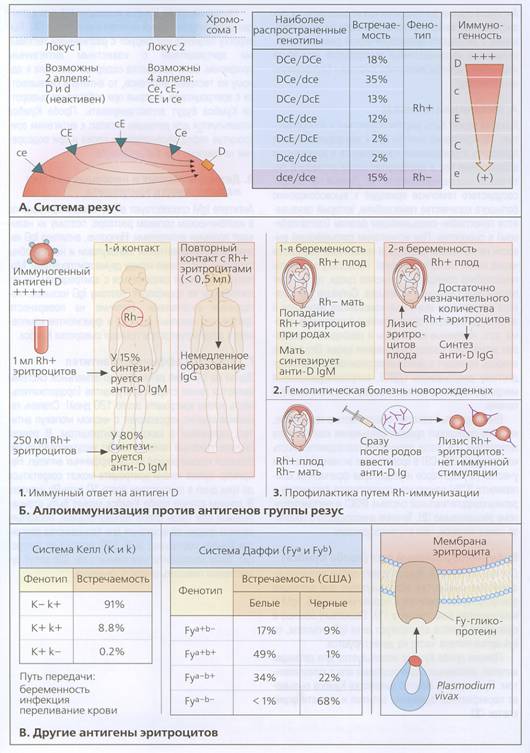
В 1949 г. Ландштейнер и Винер при иммунизации кроликов эритроцитами макака-резуса получили сыворотку, которая агглютинировала эритроциты 39 из 45 особей. При сравнении этих антител с антителами, обнаруженными Левином и Стетсоном, авторы пришли к выводу, что в обоих случаях реакция происходит с одним и тем же антигеном. В дальнейшем оказалось, что это не совсем так. В настоящее время антиген, открытый с помощью истинного анти-резус-антитела, называется LW – в честь Ландштейнера и Винера, а Rh-типирование у человека всегда проводится с сывороткой человеческого происхождения, как это было сделано в работе Левина и Стетсона. Позднее стало понятно, что именно резус-несовместимость матери и плода является причиной эритробластоза плода и гемолитической болезни новорожденных. Оказалось, что эритроциты примерно 85% всех представителей белой расы дают положительные реакции с анти-Rh-сыворотками. Семейными исследованиями было установлено, что Rh-положительные индивиды являются гомозиготами Rh/Rh или гетерозиготами Rh/rh, тогда как rh-отрицательные индивиды – это гомозиготы rh/rh.
Гипотеза Винера. В 1941 г. Винер открыл другие антитела, которые реагировали с эритроцитами 70% всех индивидов и отличались от основного фактора Rh (Rh’ по Винеру). Третий родственный фактор был открыт в 1943г.. В семейно-популяционных исследованиях выявлены все возможные комбинации этих трех факторов, причем наследовались совместно именно комбинации. Винер выдвинул гипотезу, согласно которой эти серологические "факторы" являются "агглютиногенами" и что каждый из них детерминируется одним аллелем из серии множественных аллелей одного гена.
Гипотеза Фишера. В 1943г. Фишер сформулировал более конкретную гипотезу. В то время удалось выявить еще одно антитело, анти-Hr, и Фишер, анализируя подготовленные Рейсом полные таблицы серологических данных, обнаружил, что Rh’- и Hr-факторы комплементарны. У каждого человека в крови присутствует либо антиген Rh’, либо Hr, либо оба антигена. Индивид, имеющий оба антигена, никогда не передает их вместе одному потомку, т.е. ребенок всегда получает только один антиген из двух. Для объяснения этих фактов Фишер предложил модель, согласно которой пара аллелей определяет один из двух антигенов. Эта пара была названа С/с. Аналогично была постулирована дополнительная пара аллелей D/d для исходных антигенов Rh+ и rh-, а также третья пара аллелей для уже открытого тогда третьего серологического фактора. Кроме того, чтобы согласовать генетические данные о наследовании всех трех факторов, постулировалось наличие тесного сцепления между этими тремя локусами.
Гипотеза Фишера предполагала открытие двух недостающих (комплементарных D и E) антигенов d и e. Это предсказание подтвердилось для антигена е, но не для d. По-видимому этот хромосомный район не содержит то "нечто", что приводит к образованию антител. В развитии данной гипотезы Фишер сделал важный шаг вперед. В британской популяции наиболее частыми были три класса комплексов Rh-генов. По мнению Фишера, редкие комбинации появляются вследствие изредка происходящего кроссинговера. Действительно, все четыре комбинации, относящиеся к редким классам, могут возникнуть в результате кроссинговера между более частыми комбинациями, но для CdE это не так. Для появления этой комбинации нужен двойной кроссинговер. Следовательно, гипотеза объясняет, почему комбинация CdE так редка. Возможно и другое объяснение. При каждом кроссинговере, приводящем к возникновению Cde, CDE или cde, должна возникать также и комбинация cDe. Отсюда следует, что суммарная частота первых трех комбинаций должна быть равна частоте cDe. Действительно, найденные частоты были такими: cDe – 0.0257 и Cde+cdE+CDE-0.0241 (среди негров, однако, частота cDe была выше).
Кроме того, Фишер предположил, что три указанных локуса расположены в последовательности D-C-E, поскольку комбинация cdE, которая возникает вследствие кроссинговера между локусами D и E в генотипе cDEcde, встречается намного чаще относительно этого генотипа, чем комбинация CDE относительно генотипа CDe/Cde (кроссинговер между С и Е).
После исследований Фишера было сделано много новых наблюдений. Наиболее важным для решения вопроса о порядке расположения генов было выявление комбинированных антигенов, например се. Существование этого составного антигена, по-видимому, не противоречит последовательности D-C-E, тогда как составные антигены, предполагающие тесное сцепление между D/d и E/e, не были обнаружены. Гипотеза Фишера поставила два вопроса.
1. Если вследствие кроссинговера формируются иногда редкие комбинации из более частых, то в семейных исследованиях должны обнаруживаться случаи кроссинговера. Действительно, имелось сообщение об одной такой семье: у отца с генотипом CDe/cde и матери с генотипом cde/cde было четверо детей cde/cde и трое - CDe/cde, что находится в полном соответствии с генетической теорией. Однако, шестой в порядке рождения детей имел генотип Cde/cde. Этот факт можно было бы объяснить тем, что ребенок внебрачный. Однако такое объяснение кажется малоправдоподобным, если исходить из данных по другим группам крови и сывороточным факторам, а также учитывая принадлежность этой семьи к секте с особо строгими нравами. Однако, других семей, подобных этой, обнаружено не было. Вполне вероятно, что многие исследователи не просто не станут учитывать такой атипичный случай, поскольку заподозрят здесь методическую ошибку.
2. Какова должна быть структура Rh-локуса(ов) в свете достижений молекулярной генетики? Имеются две принципиальные возможности:
А) Rh-комплекс – это один цистрон с многими мутационными сайтами. Мутационные изменения выражаются в антигенных различиях;
Б) Rh-комплекс состоит из нескольких тесно сцепленных цистронов (возможно, трех), и основные антигены отражают генетическую изменчивость по этим цистронам. В отсутствие каких-либо неопровержимых биохимических данных этот вопрос оставался неразрешенным. Определенные выводы могли сделать на основании цис-транс-теста. Поскольку составной антиген се обнаруживается только в цис-положении СЕ/се, но не в транс-положении Се/сЕ, Рейс и Сэнгер высказали гипотезу, согласно которой С/с и Е/е относятся к одному функциональному гену.
В последнее время накоплено множество фактов, которые свидетельствуют о том, что не только в структурном, но и в количественном отношении экспрессия Rh-фактора находится под строгим генетическим контролем. Розенфельд и соавт. (1973) попытались обобщить все имеющиеся данные на основе новой модели структуры Rh-локуса. Согласно этой модели, Rh-локус состоит из нескольких областей (структурных генов), несущих информацию о мембранных полипептидах. Эти области находятся под контролем общего оператора или промотора, который регулирует количественную экспрессию, возможно, благодаря нескольким операторным районам, приближенным к единственному структурному гену. Эта модель объединяет фишерскую концепцию с более поздними результатами молекулярной биологии.
В настоящее время известно, что система резус (Rh) сложна как генетически, так и фенотипически. Антигены этой системы обозначаются DdCcEe и кодируются соответствующими генами. Компонент d представляет собой молчащий аллель, наличие которого не приводит к экспрессии антигена. В соответствии с упрощенной моделью экспрессия антигенов данной группы крови контролируется двумя соседними локусами генов на хромосоме 1. В локусе 1 активным может быть молчащий ген d или ген D, кодирующий антиген D. В соседнем локусе расположены 4 аллеля: CE, Ce, cE и ce. Гены двух локусов являются кодоминантными. Другими словами, продукт гена из первого локуса экспрессируется вместе с геном из второго локуса. В результате возможны следующие комбинации: DCE, DCe, DcE, Dce, dCE, dCe, dcE, dce. Поскольку каждый человек наследует один аллель от отца, а другой от матери, то разнообразие генотипов достаточно велико. Тип людей у которых происходит экспрессия продукта гена D, называют резус-положительными (Rh+), а тип людей с генотипами, в которых антиген D потерян (активен молчащий аллель d), называют резус-отрицательными (Rh-). Антиген D по иммуногенности намного превосходит другие антигены.
Аллоимунизация против антигенов группы резус. Аллоимунизация против резусных антигенов происходит при попадании в кровь несовместимых эритроцитов. Например, это может случиться при переливании резус-отрицательному пациенту резус-положительных эритроцитов. Всего лишь 1мл Rh+ крови в 15 % случаев приводит к выработке анти-D антител класса IgM; у 80 % Rh-отрицательных индивидов выработка анти-D антител происходит после введения 250 Rh+ эритроцитов. Повторное введение даже очень малых количеств Rh+ эритроцитов может вызвать быстрое образование антител Ig G против антигена D. Важным клиническим примером служит гемолитическая болезнь новорожденных (ГБН). После родов резус-негативной матерью резус-положительного ребенка в ее организме начинают вырабатываться анти-D антитела (Ig M), что связано с соприкосновением эритроцитов плода с кровеносной системой матери (обычно в момент родов). При последующей беременности контакт даже с малыми количествами эритроцитов плода активирует выработку антител IgG, проникающих сквозь плаценту. Плод может погибнуть в утробе матери в результате тяжелой гемолитической анемии (эритробластоз плода). Продуктов расщепления гемоглобина могут попадать в головной мозг, особенно в мозговой ствол (ядерная желтуха).
Рис. 18.3. Система резус и другие системы групп крови.
Рис. 18.4. Механизмы гемолиза и детекции антител.
Для предотвращения иммунизации резус-негативным матерям в течение 72 часов после родов, амниоцентеза, выкидыша или аборта следует ввести анти-D иммуноглобулины.
Введение этих антител немедленно удалит любые эритроциты плода, которые могли попасть к матери, и тем самым предотвратит стимуляцию материнской иммунной системы и синтез антител.
Существуют и другие системы групп крови, такие как Келл, Даффи и.т.д.