 |


Принцип общего конечного пути
|
|
|
|
В структурной организации нервных сетей встречается такая ситуация, когда на одном нейроне сходятся несколько афферентных терминалей из других отделов ЦНС. Это явление принято называть конвергенцией в нейронных связях. Так, например, к одному мотонейрону подходит около 6000 колла-тералей аксонов первичных афферентов, спинальных интернейронов, нисходящих путей из стволовой части мозга и коры. Все эти терминальные окончания образуют на мотонейроне возбуждающие и тормозные синапсы и формируют своеобразную «воронку», суженная часть которой представляет общий моторный выход. Данная воронка является анатомическим образованием, определяющим один из механизмов координационной функции спинного мозга.
Суть этого механизма была раскрыта английским физиологом Ч. Шерринг-тоном, который сформулировал принцип общего конечного пути. По Ч. Шер-рингтону, количественное преобладание чувствительных и других приходящих волокон над двигательными создает неизбежное столкновение импульсов в общем конечном пути, которым являются группа мотонейронов и иннервируемые ими мышцы. В результате этого столкновения достигается торможение всех возможных степеней свободы двигательного аппарата, кроме одной, в направлении которой протекает рефлекторная реакция, вызванная максимальной стимуляцией одного из афферентных входов.
Рассмотрим случай с одновременным раздражением рецептивных полей чесательного и сгибательного рефлексов, которые реализуются идентичными группами мышц. Импульсы, идущие от этих рецептивных полей, приходят к одной и той же группе мотонейронов, и здесь в узком месте воронки за счет интеграции синаптических влияний осуществляется выбор в пользу сгибательного рефлекса, вызванного более сильным болевым раздражением. Принцип общего конечного пути, как один из принципов координации, действителен не только для спинного мозга, он применим для любого этажа ЦНС, в том числе для моторной коры.
|
|
|
Временная и пространственная суммация. Окклюзия
Конвергенция лежит в основе таких физиологических феноменов, как временная и пространственная суммация. В том случае, если два подпороговых раздражителя, приходящие к нейрону через афферентный вход, следуют друг за другом с малым временным интервалом, имеет место суммация вызванных этими раздражителями ВПСП, и суммарный ВПСП достигает порогового уровня, достаточного для генерации импульсной активности. Данный процесс способствует усилению поступающих к нейрону слабых сигналов и определяется как временная суммация. Вместе с тем синаптическая активация нейрона может осуществляться через два раздельных входа, конвергирующих на эту клетку. Одновременная стимуляция этих входов подпороговыми раздражителями также может привести к суммации ВПСП, возникающих в двух пространственно разделенных зонах клеточной мембраны. В данном случае происходит пространственная суммация, которая, так же как и временная, может вызывать длительную деполяризацию клеточной мембраны и генерацию ритмической импульсной активности на фоне этой деполяризации.
Однако возможна и такая ситуация, когда при одновременной стимуляции двух входов возбуждение нейрона и соответствующий ему рефлекторный ответ будут меньше алгебраической суммы ответов при раздельной стимуляции этих входов. При раздельной стимуляции двух входов мотонейрон б будет возбуждаться дважды: сначала совместно с нейроном а и затем совместно с нейроном в. При одновременной стимуляции двух входов нейрон б будет возбуждаться только один раз и, соответственно, рефлекторный ответ будет меньше алгебраической суммы ответов при раздельной стимуляции. Этот физиологический феномен, связанный с наличием дополнительного общего пути для двух входов, получил название окклюзии.
|
|
|
Как уже отмечалось, локальные нейронные сети могут усиливать слабые сигналы по механизму положительной обратной связи за счет циклической реверберации возбуждения в цепи нейронов. Другой возможный механизм усиления создается за счет синоптической потенциации (облегчения) при ритмических раздражениях пресинаптических входов. Потенциация выражается в увеличении амплитуды ВПСП во время (тетаническая потенциация) и после (посттетаническая потенциация) ритмического раздражения пресинапти-ческого аксона с довольно высокой частотой (100-200 имп/с).
Торможение
Координирующая функция локальных нейронных сетей помимо усиления может выражаться и в ослаблении слишком интенсивной активности нейронов за счет их торможения. Торможение, как особый нервный процесс, характеризуется отсутствием способности к активному распространению по нервной клетке и может быть представлено двумя, формами — первичным и вторичным торможением. Первичное торможение обусловлено наличием специфических тормозных структур и развивается первично без предвательного возбуждения. Примером первичного торможения является так называемое реципрокное торможение мыщц-антагонистов, обнаруженное в спинальных рефлекторных дугах. Суть явлений состоит в том, что если активируются приорецепторы мышцы-сгибателя, то через первичные афференты одновременно возбуждают мотонейрон данной мыпш сгибателя и через коллатераль афферентного волокна — тормозный вставочный нейрон. Возбуждение вставочного нейрона приводит к постсинаптическому торможению мотонейрона антагонистичной мышцы-разгибателя, на теле которого аксон тормозного интернейрона формирует специализированные тормозные синапсы. Реципрокное торможение играет важную роль в автоматической координации двигательных актов.
Торможение по принципу отрицательной обратной связи встречается только на выходе, но и на входе моторных центров спинного мозга. Явление подобного рода описано в моносинаптических соединениях афферентных волокон со спинальными мотонейронами, торможение которых при данной ситуации не связано с изменениями в постсинаптической мембране. Последнее обстоятельство позволило определить эту форму торможения как пресинаптическое. Оно обусловлено наличием вставочных тормозных нейронов, к которым подходят коллатерали афферентных волокон. В свою очередь, вставочные нейроны формируют аксо-аксональные синапсы на афферентных терминалях, являющихся пресинаптическими по отношению к мотонейронам.
|
|
|
СЕДЬМОЙ вопрос.
В ЦНС различают более древние сегментарные и эволюционно более молодые надсегментарные отделы нервной системы. К сегментарным отделам относят спинной, продолговатый и средний мозг, участки которых регулируют функции отдельных участков тела, лежащих на том же уровне. Надсегментарные отделы: промежуточный мозг, мозжечок и кора больших полушарий - не имеют непосредственных связей с органами тела, а управляют их деятельностью через нижележащие сегментарные отделы.
Спинной мозг. Спинной мозг является низшим и наиболее древним отделом ЦНС. В составе серого вещества спинного мозга человека насчитывают около 13,5 млн нервных клеток. Из них основную массу (97%) представляют промежуточные клетки (вставочные, или интернейроны), которые обеспечивают сложные процессы координации внутри спинного мозга. Среди мотонейронов спинного мозга различают крупные клетки - альфа-мотонейроны и мелкие - гамма-мотонейроны. От альфа-мотонейронов отходят наиболее толстые и быстропроводящие волокна двигательных нервов, вызывающие сокращения скелетных мышечных волокон. Тонкие волокна гамма-мотонейронов не вызывают сокращения мышц. Они подходят к про-приорецепторам - мышечным веретенам и регулируют чувствительность этих рецепторов, информирующих мозг о выполнении движений.
Рефлексы спинного мозга можно подразделить на двигательные, осуществляемые альфа-мотонейронами передних рогов, и вегетативные, осуществляемые эфферентными клетками боковых рогов. Мотонейроны спинного мозга иннервируют все скелетные мышцы (за исключением мышц лица). Спинной мозг осуществляет элементарные двигательные рефлексы: сгибательные и разгибательные, ритмические, шагательные, возникающие при раздражении кожи или проприорецепторов мышц и сухожилий, а также посылает постоянную импульсацию к мышцам, поддерживая их напряжение - мышечный тонус. Специальные мотонейроны иннервируют дыхательную мускулатуру - межреберные мышцы и диафрагму - и обеспечивают дыхательные движения. Вегетативные нейроны иннервируют все внутренние органы (сердце, сосуды, потовые железы, железы внутренней секреции, пищеварительный тракт, мочеполовую систему) и осуществляют рефлексы, регулирующие их деятельность.
|
|
|
Проводниковая функция спинного мозга связана с передачей в вышележащие отделы нервной системы получаемого с периферии потока информации и с проведением импульсов, идущих из головного мозга в спинной.
Продолговатый мозг и варолиев мост. Продолговатый мозг и варо-лиев мост являются частью ствола мозга. Здесь находится большая группа черепных нервов (от V до XII пары), иннервирующих кожу, слизистые оболочки, мускулатуру головы и ряд внутренних органов (сердце, легкие, печень). Тут же находятся центры многих пищеварительных рефлексов: жевания, глотания, движений желудка и части кишечника, выделения пищеварительных соков, а также центры некоторых защитных рефлексов (чихания, кашля, мигания, слезоотделения, рвоты) и центры водно-солевого и сахарного обмена. На дне IV желудочка в продолговатом мозге находится жизненно важный дыхательный центр. В непосредственной близости расположен сердечно-сосудистый центр. Его крупные клетки регулируют деятельность сердца и просвет сосудов.
Продолговатый мозг играет важную роль в осуществлении двигательных актов и в регуляции тонуса скелетных мышц, повышая тонус мышц-разгибателей. Он, в частности, принимает участие в осуществлении установочных рефлексов позы (шейных, лабиринтных).
Через продолговатый мозг проходят восходящие пути - слуховой, вестибулярный, проприоцептивный и тактильной чувствительности.
Средний мозг. В состав среднего мозга входят четверохолмия, черная субстанция и красные ядра. В передних буграх четверохолмия находятся зрительные подкорковые центры, а в задних - слуховые. Средний мозг участвует в регуляции движений глаз, осуществляет зрачковый рефлекс (расширение зрачков в темноте и сужение их на свету).
Четверохолмия выполняют ряд реакций, являющихся компонентами ориентировочного рефлекса. В ответ на внезапное раздражение происходит поворот головы и глаз в сторону раздражителя. Этот рефлекс (по И.П. Павлову - рефлекс «Что такое?») необходим для подготовки организма к своевременной реакции на любое новое воздействие.
|
|
|
Черная субстанция среднего мозга имеет отношение к рефлексам жевания и глотания, участвует в регуляции тонуса мышц (особенно при выполнении мелких движений пальцами рук) и в организации содружественных двигательных реакций.
Красное ядро среднего мозга выполняет моторные функции: регулирует тонус скелетных мышц, вызывая усиление тонуса мышц-сгибателей.
Оказывая значительное влияние на тонус скелетных мышц, средний мозг принимает участие в ряде установочных рефлексов поддержания позы (выпрямительных - установке тела теменем вверх и др.).
Промежуточный мозг. В состав промежуточного мозга входят тала-мус (зрительные бугры) и гипоталамус (подбугорье).
Через таламус проходят все афферентные пути (за исключением обонятельных), которые направляются в соответствующие воспринимающие области коры (слуховые, зрительные и пр.). Ядра таламуса подразделяются на специфические и неспецифические. К специфическим относят переключательные (релейные) ядра и ассоциативные. Через переключательные ядра таламуса передаются афферентные влияния от всех рецепторов тела. Ассоциативные ядра получают импульсы от переключательных ядер и обеспечивают их взаимодействие, т.е. осуществляют их подкорковую интеграцию. Помимо этих ядер в таламусе имеются неспецифические ядра, которые оказывают как активирующие, так и тормозящие влияния на небольшие области коры.
Благодаря обширным связям таламус играет важнейшую роль в жизнедеятельности организма. Импульсы, идущие от таламуса в кору, изменяют состояние корковых нейронов и регулируют ритм корковой активности. При непосредственном участии таламуса происходит образование условных рефлексов и выработка двигательных навыков, формирование эмоций человека, его мимики. Таламус играет большую роль в возникновении ощущений, в частности ощущения боли. С его деятельностью связывают регуляцию биоритмов в жизни человека (суточных, сезонных и др.).
Гипоталамус является высшим подкорковым центром регуляции вегетативных функций. Здесь расположены вегетативные центры, регулирующие обмен веществ в организме, обеспечивающие поддержание постоянства температуры тела (у теплокровных) и нормального уровня кровяного давления, поддерживающие водный баланс, регулирующие чувство голода и насыщения. Раздражение задних ядер гипоталамуса вызывает усиление симпатических влияний, а передних - парасимпатические эффекты.
Благодаря тесной связи гипоталамуса с гипофизом (гипоталамо-гипофизарная система) осуществляется контроль за деятельностью желез внутренней секреции. Вегетативные и гормональные реакции, регулируемые гипоталамусом, являются компонентами эмоциональных и двигательных реакций человека. Структуры гипоталамуса связаны также с регуляцией состояний бодрствования и сна.
Неспецифическая система мозга. Неспецифическая система занимает срединную часть ствола мозга. Она не связана с анализом какой-либо специфической чувствительности или с выполнением определенных рефлекторных реакций. Импульсы в эту систему поступают через боковые ответвления от всех специфических путей, в результате чего обеспечивается их обширное взаимодействие.
Для неспецифической системы характерно расположение нейронов в виде диффузной сети, обилие и разнообразие их отростков. В связи с этим она и получила название сетевидного образования, или ретикулярной формации.
Различают два типа влияния неспецифической системы на работу других нервных центров - активирующее и тормозящее. Оба типа этих влияний могут быть восходящими (к вышележащим центрам) и нисходящими (к нижележащим центрам). Они служат для регулирования функционального состояния мозга, уровня бодрствования и регуляции позно-тонических и фазных реакций скелетных мышц.
Мозжечок. Мозжечок - это надсегментарное образование, не имеющее непосредственных связей с исполнительными аппаратами. Мозжечок состоит из непарного образования - червя и парных полушарий. Основными нейронами коры мозжечка являются многочисленные клетки Пуркине. Благодаря обширным связям (на каждой клетке оканчивается до 200 000 синапсов) в них происходит интеграция самых различных сенсорных влияний, в первую очередь проприоцептивных, тактильных и вестибулярных. Представительство разных периферических рецепторов в коре мозжечка имеет сома-тотопическую организацию (от греч. somatos - тело, topos - место), т.е. отражает порядок их расположения в теле человека. Кроме того, этот порядок расположения соответствует такому же порядку расположения представительства участков тела в коре больших полушарий, что облегчает обмен информацией между корой и мозжечком и обеспечивает их совместную деятельность в управлении поведением человека. Правильная геометрическая организация нейронов мозжечка обуславливает его значение в отсчете времени и четком поддержании темпа циклических движений.
Основной функцией мозжечка является регуляция позно-тонических реакций и координация двигательной деятельности.
По анатомическим особенностям (связям коры мозжечка с его ядрами) и функциональному значению мозжечок подразделяют на три продольные зоны: внутреннюю, или медиальную, - кору червя, функцией которой является регуляция тонуса скелетных мышц, поддержание позы и равновесия тела; промежуточную - среднюю часть коры полушарий мозжечка, функция которой - согласование позных реакций с движениями, а также коррекция ошибок; боковую, или латеральную, - кору полушарий мозжечка, которая совместно с промежуточным мозгом и корой больших полушарий участвует в программировании быстрых баллистических движений (бросков, ударов, прыжков и др.).
Базальные ядра. К базальным ядрам относят полосатое ядро, состоящее из хвостатого ядра и скорлупы, и бледное ядро, а в настоящее время причисляют также миндалевидное тело (относящееся к вегетативным центрам лимбической системы) и черную субстанцию среднего мозга.
Афферентные влияния приходят к базальным ядрам от рецепторов тела через таламус и от всех областей коры больших полушарий. Они поступают в полосатое тело. Эфферентные влияния от него направляются к бледному ядру и далее к стволовым центрам экстрапирамидной системы, а также через таламус обратно к коре.
Базальные ядра участвуют в образовании условных рефлексов и осуществлении сложных безусловных рефлексов (оборонительные, пищедобы-вательные и др.). Они обеспечивают необходимое положение тела во время физической работы, а также протекание автоматических ритмических движений (древних автоматизмов).
Бледное ядро выполняет основную моторную функцию, а полосатое тело регулирует его активность. В настоящее время выявлено значение хвостатого ядра в контроле сложных психических процессов - внимания, памяти, обнаружения ошибок.
ВОСЬМОЙ вопрос.
У высших млекопитающих - животных и человека - ведущим отделом ЦНС является кора больших полушарий.
Корковые нейроны. Кора представляет собой слой серого вещества толщиной 2-3 мм, содержащий в среднем около 14 млрд нервных клеток. Для нее характерно обилие межнейронных связей.
Основными типами корковых клеток являются звездчатые и пирамидные нейроны. Звездчатые нейроны связаны с процессами восприятия раздражений и объединением деятельности различных пирамидных нейронов. Пирамидные нейроны осуществляют эфферентную функцию коры (преимущественно через пирамидный тракт) и внутрикорковые процессы взаимодействия между удаленными друг от друга нейронами. Наиболее крупные пирамидные клетки - гигантские пирамиды Беца - находятся в передней центральной извилине (моторной зоне коры).
Функциональной единицей коры является вертикальная колонка взаимосвязанных нейронов. Вытянутые по вертикали крупные пирамидные клетки с расположенными над ними и под ними нейронами образуют функциональные объединения нейронов. Все нейроны вертикальной колонки отвечают на одно и то же афферентное раздражение (от одних и тех же рецепторов) одинаковой реакцией и совместно формируют эфферентные ответы пирамидных нейронов.
 Схема корковой функциональной единицы — вертикальной колонки нейронов
Схема корковой функциональной единицы — вертикальной колонки нейронов
1,2 - пирамидные нейроны; 3, 4 - возвратные коллатерали аксонов; 5 – эфферентный выход; 6, 7 - афферентные входы; 8 – интернейрон
По мере надобности вертикальные колонки могут объединяться в более крупные образования, обеспечивая комбинированные реакции. Функциональное значение различных корковых полей. По особенностям строения и функциональному значению отдельных корковых участков вся кора подразделяется на три основные группы полей - первичные, вторичные и третичные.
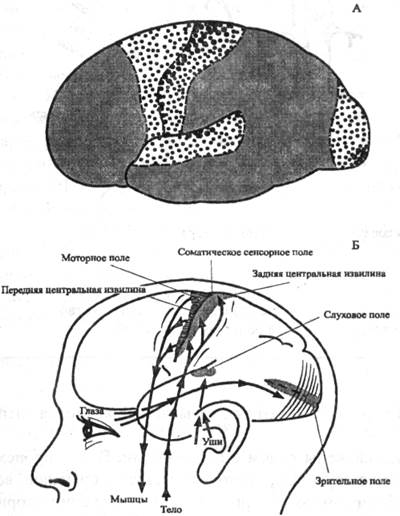 Первичные поля связаны с органами чувств и органами движения на периферии. Они обеспечивают возникновение ощущений. К ним относятся, например, поле болевой и мышечно-суставной чувствительности в задней центральной извилине коры, зрительное поле в затылочной области, слуховое поле в височной области и моторное поле в передней центральной извилине. В первичных полях находятся высокоспециализированные клетки-определители, или детекторы, избирательно реагирующие только на определенные раздражения.
Первичные поля связаны с органами чувств и органами движения на периферии. Они обеспечивают возникновение ощущений. К ним относятся, например, поле болевой и мышечно-суставной чувствительности в задней центральной извилине коры, зрительное поле в затылочной области, слуховое поле в височной области и моторное поле в передней центральной извилине. В первичных полях находятся высокоспециализированные клетки-определители, или детекторы, избирательно реагирующие только на определенные раздражения.
Первичные, вторичные и третичные поля коры больших полушарий
На А: крупные точки -первичные поля, средние - вторичные поля, мелкие (серый фон) - третичные поля. На Б: первичные (проекционные) поля коры больших полушарий
Например, в зрительной коре имеются нейроны-детекторы, возбуждающиеся только при включении или выключении света, чувствительные лишь к определенной его интенсивности, к конкретным интервалам светового воздействия, к определенной длине волны и т.д.
 При разрушении первичных полей коры возникают так называемые корковая слепота, корковая глухота и т.п. Вторичные поля расположены рядом с первичными. В них происходит осмысливание и узнавание звуковых, световых и других сигналов, возникают сложные формы обобщенного восприятия. При поражении вторичных полей сохраняется способность видеть предметы, слышать звуки, но человек их не узнает, не помнит значения.
При разрушении первичных полей коры возникают так называемые корковая слепота, корковая глухота и т.п. Вторичные поля расположены рядом с первичными. В них происходит осмысливание и узнавание звуковых, световых и других сигналов, возникают сложные формы обобщенного восприятия. При поражении вторичных полей сохраняется способность видеть предметы, слышать звуки, но человек их не узнает, не помнит значения.
Сенсорное (слева) и моторное (справа) представительство различных частей тела в коре больших полушарий
Третичные поля развиты только у человека. Это ассоциативные области коры, обеспечивающие высшие формы анализа и синтеза и формирующие целенаправленную поведенческую деятельность человека. Третичные поля находятся в задней половине коры - между теменными, затылочными и височными областями - и в передней половине - в передних частях лобных областей. Их роль особенно велика в организации согласованной работы обоих полушарий. Третичные поля созревают у человека позже других корковых полей и раньше других деградируют при старении организма.
Функцией задних третичных полей (главным образом, нижнетеменных областей коры) является прием, переработка и хранение информации. Они формируют представление о схеме тела и схеме пространства, обеспечивая пространственную ориентацию движений. Передние третичные поля (переднелобные области) выполняют общую регуляцию сложных форм поведения человека, формируя намерения и планы, программы произвольных движений и контроль за их выполнением. Развитие третичных полей у человека связывают с функцией речи. Мышление(внутренняя речь) возможно только при совместной деятельности различных сенсорных систем, объединение информации от которых происходит в третичных полях. При врожденном недоразвитии третичных полей человек не в состоянии овладеть ре- • чью (произносит лишь бессмысленные звуки) и даже простейшими двигательными навыками (не может одеваться, пользоваться орудиями труда и т.п.).
Парная деятельность и доминирование полушарий. Обработка информации осуществляется в результате парной деятельности обоих полушарий головного мозга. Однако, как правило, одно из полушарий является ведущим - доминантным. У большинства людей с ведущей правой рукой (правшей) доминантным является левое полушарие, а соподчиненным (субдоминантным) - правое полушарие.
Левое полушарие по сравнению с правым имеет более тонкое нейронное строение, большее богатство взаимосвязей нейронов, более концентрированное представительство функций и лучшие условия кровоснабжения. В левом доминантном полушарии находится моторный центр речи (центр Брока), обеспечивающий речевую деятельность, и сенсорный центр речи, осуществляющий понимание слов. Левое полушарие специализировано на тонком сенсомоторном контроле за движениями рук.
Функциональная асимметрия обнаруживается у человека в отношении не только моторных функций (моторная асимметрия), но и сенсорных (сенсорная асимметрия). Как правило, у человека имеется «ведущий глаз» и «ведущее ухо», сигналы от которых являются главенствующими при восприятии. Однако проблема функциональной асимметрии довольно сложна. Например, у человека-правши может быть ведущим левый глаз или левое ухо. В каждом полушарии могут быть представлены функции не только противоположной, но и одноименной стороны тела. В результате этого обеспечивается возможность замещения одного полушария другим в случае его повреждения, а также создается структурная основа для переменного доминирования полушарий при управлении движениями.
Специализация полушарий проявляется и в отношении психических функций {психическая асимметрия). Для левого полушария характерны аналитические процессы, последовательная обработка информации, в том числе с помощью речи, абстрактное мышление, оценка временных отношений, предвосхищение будущих событий, успешное решение вербально-логи-ческих задач. В правом полушарии информация обрабатывается целостно, синтетически (без расчленения на детали), с учетом прошлого опыта и безучастия речи, преобладает предметное мышление. Эти особенности позволяют связывать с правым полушарием восприятие пространственных признаков и решение зрительно-пространственных задач.
Электрическая активность коры больших полушарий. Изменение функционального состояния коры отражаются в записи ее электрической активности - электроэнцефалограммы (ЭЭГ). Современные электроэнцефалографы усиливают потенциалы мозга в 2-3 млн раз и дают возможность исследовать ЭЭГ от многих точек коры одновременно, т.е. изучать системные процессы.
Различают определенные диапазоны частот, называемые ритмами ЭЭГ, в состоянии относительного покоя чаще всего регистрируется альфа-ритм (8-13 колебаний в 1 с); в состоянии активного внимания - бета-ритм (14 колебаний в 1 с и выше); при засыпании, некоторых эмоциональных состояниях - тета-ритм (4-7 колебаний в 1 с); при глубоком сне, потере сознания, наркозе - дельта-ритм (1-3 колебания в 1 с).
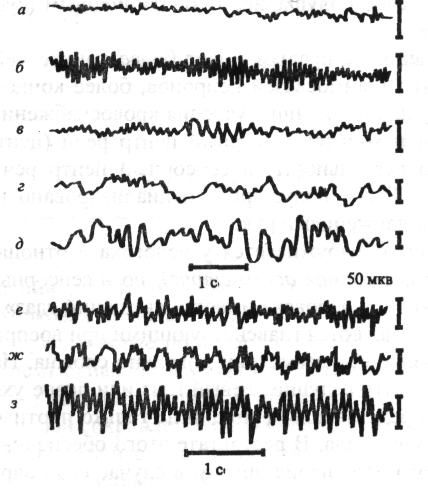 Электроэнцефалограмма затылочной (а - д) и моторной (е - з) областей коры больших полушарий человека при различных состояниях и во время мышечной работы:
Электроэнцефалограмма затылочной (а - д) и моторной (е - з) областей коры больших полушарий человека при различных состояниях и во время мышечной работы:
а - активное состояние, глаза открыты (бета-ритм); б - покой, глаза закрыты (альфа-ритм); в - дремота (тета-ритм); г - засыпание; д - глубокий сон (дельта-ритм); е - непривычная или тяжелая работа -асинхронная частая активность (явление десинхронизации); ж -циклические движения - медленные потенциалы в темпе движения («меченые ритмы» ЭЭГ); з - выполнение освоенного движения -появление альфа-ритма
Помимо фоновой активности в ЭЭГ выделяют отдельные потенциалы, связанные с какими-либо событиями: вызванные потенциалы, возникающие в ответ на внешние раздражения (слуховые, зрительные и др.); потенциалы, отражающие мозговые процессы при подготовке, осуществлении и окончании отдельных двигательных актов - «волна ожидания», или условная негативная волна: премоторные, моторные, финальные потенциалы и др. Кроме того, регистрируют сверхмедленные колебания длительностью от нескольких секунд до десятков минут (так называемые «омега-потенциалы» и др.), которые отражают биохимические процессы регуляции функций и психической деятельности.
ДЕВЯТЫЙ вопрос.
Под лимбической системой понимают ряд корковых и подкорковых структур, функции которых связаны с организацией мотивационно-эмоциональных реакций, процессами памяти и обучения.
Корковые отделы лимбической системы, представляющие ее высший отдел, находятся на нижних и внутренних поверхностях больших полушарий (участки лобной коры, поясная извилина, или лимбическая кора, гиппокамп и др.). К подкорковым структурам лимбической системы относят гипоталамус, некоторые ядра таламуса, среднего мозга и ретикулярной формации. Между всеми этими образованиями имеются тесные прямые и обратные связи, образующие так называемое лимбическое кольцо.
Лимбическая система участвует в самых разнообразных проявлениях деятельности организма: в регуляции пищевого и питьевого поведения, цикла сон-бодрствование, в процессах формирования памятного следа (отложения и извлечения из памяти), в развитии агрессивно-оборонительных реакций, обеспечивая избирательный характер поведения. Она формирует положительные и отрицательные эмоции со всеми двигательными и гормональными их компонентами. Исследование различных участков лимбической системы выявило наличие центров удовольствия, формирующих положительные эмоции, и неудовольствия, формирующих отрицательные эмоции. Изолированное раздражение таких точек в глубоких структурах мозга человека вызывало появление чувства «беспричинной радости», «беспредметной тоски», «безотчетного страха».
ДЕСЯТЫЙ вопрос.
Все функции организма условно можно разделить на соматические, или анимальные (животные), связанные с восприятием внешней информации и деятельностью мышц, и вегетативные (растительные), связанные с деятельностью внутренних органов: процессы дыхания, кровообращения, пищеварения, выделения, обмена веществ, роста и размножения.
Функциональная организация вегетативной нервной системы. Вегетативной нервной системой называют совокупность эфферентных нервных клеток спинного и головного мозга, а также клеток особых узлов (ганглиев), иннервирующих внутренние органы. Раздражения различных рецепторов тела могут вызвать изменения как соматических, так и вегетативных функций, поскольку афферентные и центральные отделы этих рефлекторных дуг общие. Они различаются лишь своими эфферентными отделами. Характерной особенностью эфферентных путей, входящих в рефлекторные дуги вегетативных рефлексов, является их двухнейронное строение (один нейрон находится в ЦНС, другой - в ганглиях или в иннервируемом органе).
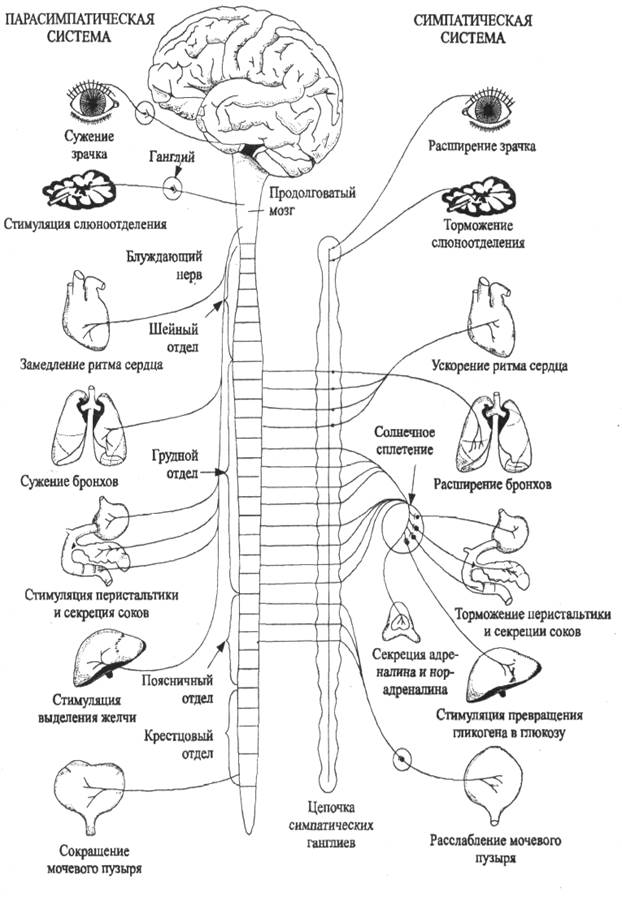
Вегетативная нервная система подразделяется на два отдела - симпатический и парасимпатический.
Эфферентные пути симпатической нервной системы начинаются в грудном и поясничном отделах спинного мозга от нейронов его боковых рогов. Передача возбуждения с предузловых симпатических волокон на после-узловые происходит с участием медиатора ацетилхолина, а с послеузловых волокон на иннервируемые органы - с участием медиатора норадреналина. Исключением являются волокна, иннервирующие потовые железы и расширяющие сосуды скелетных мышц, где возбуждение передается с помощью ацетилхолина.
Эфферентные пути парасимпатической нервной системы начинаются в головном мозге - от некоторых ядер среднего и продолговатого мозга - и в спинном мозге - от нейронов крестцового отдела. Проведение возбуждения в синапсах парасимпатического пути происходит с участием медиатора ацетилхолина. Второй нейрон находится в иннервируемом органе или вблизи него.
Высшим регулятором вегетативных функций является гипоталамус, который действует совместно с ретикулярной формацией и лимбической системой, под контролем коры больших полушарий. Кроме того, нейроны, расположенные в самих органах или в симпатических узлах, могут осуществлять собственные рефлекторные реакции без участия ЦНС - «периферические рефлексы».
Функции симпатической нервной системы. С участием симпатической нервной системы протекают многие важные рефлексы в организме, направленные на обеспечение его деятельного состояния, в том числе - его двигательной деятельности. К ним относятся рефлексы расширения бронхов, учащения и усиления сердечных сокращений, выброс депонированной крови из печени и селезенки, расщепление гликогена до глюкозы в печени (мобилизация углеводных источников энергии), усиление деятельности желез внутренней секреции и потовых желез. Симпатическая нервная система снижает деятельность ряда внутренних органов: в результате сужения сосудов в почках уменьшаются процессы мочеобразования, угнетается секреторная и моторная деятельность органов желудочно-кишечного тракта; предотвращается акт мочеиспускания - расслабляются мышцы стенки мочевого пузыря и сокращается его сфинктер.
Повышенная активность организма сопровождается симпатическим рефлексом расширения зрачка. Огромное значение для двигательной деятельности организма имеет трофическое влияние симпатических нервов на скелетные мышцы, улучшающее в них обмен веществ и снимающее утомление.
Вегетативная нервная система
Симпатический отдел нервной системы не только повышает уровень функционирования организма, но и мобилизует его скрытые функциональные резервы, активирует деятельность мозга, повышает защитные реакции (иммунные реакции, барьерные механизмы и др.), запускает гормональные реакции. Особенное значение имеет симпатическая нервная система при развитии стрессовых состояний, в наиболее сложных условиях жизнедеятельности. Важна роль симпатических влияний в процессах приспособления (адаптации) организма к напряженной работе в различных условиях внешней среды. Эта функция называется адаптационно-трофической.
Функции парасимпатической нервной системы. Парасимпатическая нервная система осуществляет сужение бронхов, замедление и ослабление сердечных сокращений, пополнение энергоресурсов (синтез гликогена в печени и усиление процессов пищеварения), усиление процессов мочеобра-зования в почках и обеспечение акта мочеиспускания (сокращение мышц мочевого пузыря и расслабление его сфинктера) и др. Парасимпатическая нервная система оказывает преимущественно пусковые влияния: сужение зрачков, бронхов, включение деятельности пищеварительных желез и т.п.
Деятельность парасимпатического отдела вегетативной нервной системы направлена на текущую регуляцию функционального состояния, на поддержание постоянства внутренней среды - гомеостаза. Парасимпатический отдел обеспечивает восстановление различных физиологических показателей, резко измененных после напряженной мышечной работы, пополнение израсходованных энергоресурсов. Медиатор парасимпатической системы - ацетилхолин, снижая чувствительность адренорецепторов к действию адреналина и норадреналина, оказывает определенное антистрессорное влияние.
Вегетативные рефлексы. Через вегетативные симпатические и парасимпатические пути ЦНС осуществляет некоторые вегетативные рефлексы, начинающиеся с различных рецепторов внешней и внутренней среды: висцеро-висцеральные (с внутренних органов на внутренние органы - например дыхательно-сердечный рефлекс); дермо-висцеральные (с кожных покровов - изменение деятельности внутренних органов при раздражении активных точек кожи, например, иглоукалыванием, точечным массажем); с рецепторов глазного яблока - глазо-сердечный рефлекс Аншера (урежение сердцебиений при надавливании на глазные яблоки - парасимпатический эффект); моторно-висцеральные и др. Они используются для оценки функционального состояния организма, и особенно состояния вегетативной нервной системы. По ним судят об усилении влияния симпатического или парасимпатического ее отдела.
|
|
|
