 |


Реакция хищника на увеличение численности жертв
|
|
|
|
Ограничение роста популяции жертв хищниками возможно, если в ответ на увеличение численности жертв будет возрастать удельное (т. е. приходящееся в среднем на одну особь жертв) потребление их хищниками. Подобное усиление пресса хищников в принципе может происходить, во-первых, за счет возрастания среднего рациона одной особи хищника, а во-вторых, за счет увеличения численности популяции хищника. Очевидно, первый эффект, называемый функциональной реакцией, — краткосрочный, а второй, называемый численной реакцией, — долгосрочный.

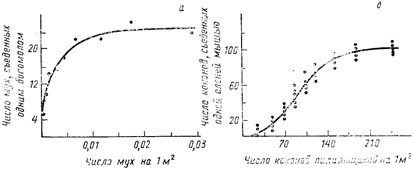 Функциональная реакция хищника на увеличение численности жертв изучена довольно хорошо. В 50-х гг., изучая экспериментальными методами питание рыб, В. С. Ивлев показал, что индивидуальный рацион (т. е. количество пищи, потребленной за единицу времени) животного при увеличении плотности предлагаемых ему кормовых объектов растет, а затем выходит на плато. Математическая модель, предложенная В. С. Ивлевым (1955) для описания зависимости величины рациона от количества предложенной пищи, в дальнейшем стала широко применяться при изучении питания самых разных (преимущественно, правда, водных) животных. Немного позже функциональная реакция хищника была детально исследована канадским экологом К. Холдингом (Holling, 1965), показавшим, что рост потребления хищником своей добычи по мере увеличения ее количества может происходить по-разному. Так, например, у богомола Hierodula crassa при кормлении его мухами (Musca domestica) число съеденных жертв сначала быстро увеличивалось, а затем снижалось и выходило на плато (рис. 41, а). В то же время у оленьей мыши Peromyscus maniculatus, которой в качестве жертв предлагали коконы соснового пилильщика Neodiprion sertifer (насекомого из отряда перепончатокрылых), потребление жертв по мере увеличения их количества росло сначала медленно, потом быстрее, а затем, замедляясь, выходило на плато, т. е. описывалось S-образной кривой (рис. 41, б). Различия между этими кривыми, видимо, отражают разные эволюционно сложившиеся стратегии добывания пищи.
Функциональная реакция хищника на увеличение численности жертв изучена довольно хорошо. В 50-х гг., изучая экспериментальными методами питание рыб, В. С. Ивлев показал, что индивидуальный рацион (т. е. количество пищи, потребленной за единицу времени) животного при увеличении плотности предлагаемых ему кормовых объектов растет, а затем выходит на плато. Математическая модель, предложенная В. С. Ивлевым (1955) для описания зависимости величины рациона от количества предложенной пищи, в дальнейшем стала широко применяться при изучении питания самых разных (преимущественно, правда, водных) животных. Немного позже функциональная реакция хищника была детально исследована канадским экологом К. Холдингом (Holling, 1965), показавшим, что рост потребления хищником своей добычи по мере увеличения ее количества может происходить по-разному. Так, например, у богомола Hierodula crassa при кормлении его мухами (Musca domestica) число съеденных жертв сначала быстро увеличивалось, а затем снижалось и выходило на плато (рис. 41, а). В то же время у оленьей мыши Peromyscus maniculatus, которой в качестве жертв предлагали коконы соснового пилильщика Neodiprion sertifer (насекомого из отряда перепончатокрылых), потребление жертв по мере увеличения их количества росло сначала медленно, потом быстрее, а затем, замедляясь, выходило на плато, т. е. описывалось S-образной кривой (рис. 41, б). Различия между этими кривыми, видимо, отражают разные эволюционно сложившиеся стратегии добывания пищи.
|
|
|
Беспозвоночные хищники, как правило, очень четко реагируют на строго определенные стимулы, специфически связанные с их жертвами. При увеличении плотности жертв хищник начинает поедать их больше, поскольку они чаще ему встречаются. Таким образом, сначала рост рациона хищника пропорционален частоте его встреч с жертвами. Однако вскоре рост этот начинает замедляться, и кривая функциональной реакции выходит на плато. Происходит это по двум причинам. Во-первых, по мере того как хищник наедается, повышается порог возбуждения, при котором наблюдается его реакция на жертву. Так в опытах К. Холдинга наевшийся богомол реагировал на мух, находящихся на меньшем расстоянии, чем голодный. Во-вторых, сам процесс поимки добычи и поедания ее требует определенного времени, и хищник не может приступить к нападению на следующую жертву, пока он не съел предыдущую.
Позвоночные хищники, обладая высоко развитой нервной системой, могут реагировать на более широкий круг раздражителей, но порог возбуждения их при этом выше (т. е. хищник как бы не отвлекается по пустякам). Поэтому при незначительном увеличении плотности жертв интенсивность потребления их позвоночными хищниками возрастает медленно (заметим также, что хищнику невыгодно расходовать много энергии на малочисленную жертву). Однако при высокой численности жертв какого-то определенного вида позвоночные хищники могут быстро обучиться или «настроиться» на преимущественное потребление именно этого конкретного вида[43]. Быстрому росту рациона соответствует средний, наиболее крутой участок S-образной кривой. В деталям форма кривой функциональной реакции может сильно варьировать от одного вида хищников к другому, а также в пределах одного вида в зависимости от возраста и физиологического состояния особей. Различие между функциональными реакциями беспозвоночных и позвоночных хищников не жесткое, а иногда S-образная кривая обнаруживается и у беспозвоночных хищников.
|
|
|
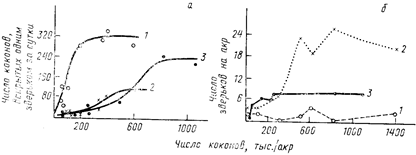 Степень проявления функциональной реакции (т. е. величина, показывающая, во сколько раз возрастает рацион при избытке пищи) у разных видов может сильно варьироваться. Так, наблюдения в природной обстановке показали (Holling, 1959), что при увеличении числа коконов соснового пилильщика[44] среднее (приходящееся на одного хищника) потребление их короткохвостой бурозубкой (Blarina brevicauda) резко возрастает, а потребление обыкновенной бурозубкой (Sorex cinereus) остается на том же уровне (рис. 42, а).
Степень проявления функциональной реакции (т. е. величина, показывающая, во сколько раз возрастает рацион при избытке пищи) у разных видов может сильно варьироваться. Так, наблюдения в природной обстановке показали (Holling, 1959), что при увеличении числа коконов соснового пилильщика[44] среднее (приходящееся на одного хищника) потребление их короткохвостой бурозубкой (Blarina brevicauda) резко возрастает, а потребление обыкновенной бурозубкой (Sorex cinereus) остается на том же уровне (рис. 42, а).
 В качестве теоретической базы их на протяжении последних двух десятилетий нередко служила концепция, известная как «теория оптимального добывания пищи» (optimal foraging theory — англ.). В основе ее лежит, в общем, очевидное предположение о том, что в ходе эволюции максимизируется количество энергии, получаемое животным за определенное время, или минимизируется время, потраченное на добывание пищи.
В качестве теоретической базы их на протяжении последних двух десятилетий нередко служила концепция, известная как «теория оптимального добывания пищи» (optimal foraging theory — англ.). В основе ее лежит, в общем, очевидное предположение о том, что в ходе эволюции максимизируется количество энергии, получаемое животным за определенное время, или минимизируется время, потраченное на добывание пищи.
Численная реакция хищника на увеличение плотности популяции жертв также сильно изменяется от одного вида к другому. В частности, в той же работе К. Холдинга показано, что при увеличении числа коконов соснового пилильщика заметно возрастает численность обыкновенной бурозубки, тогда как численность другого вида землероек — короткохвостой бурозубки — остается почти неизменной (рис. 42, б). Иначе говоря, наблюдаемая ситуация обратная той, что отмечалась при сопоставлении функциональных реакций этих же видов. Что же касается реакции третьего изученного К. Холдингом вида — оленьей мыши (Peromyscus maniculatus), то его функциональная и численная реакции оказались промежуточными между отмеченными для обыкновенной и короткохвостой бурозубок.
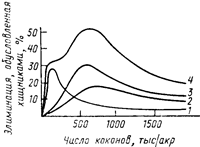
Зная функциональную и численную реакцию каждого из этих видов, оказалось возможным оценить их суммарный эффект на популяцию жертв. Выяснилось, что при незначительном подъеме плотности жертв существенная роль в ограничении их популяций принадлежит короткохвостой бурозубке, а при более значительном — обыкновенной бурозубке и оленьей мыши (рис. 43). Максимальный удельный (в расчете на одну особь жертвы) пресс хищников достигается при некоторой средней плотности жертв. При дальнейшем же возрастании плотности жертв доля популяции, изымаемая хищником, снижается, и фактически популяция не контролируется хищниками.
|
|
|
Если выход на плато кривой функциональной реакции понятен (не может же один человек, даже очень голодный, съесть за день слона), то причины остановки роста численности хищников при наличии избытка доступных жертв менее ясны. По-видимому, в самом общем виде можно предполагать, что численность популяции хищников начинает лимитироваться другими фактором, например поведенческими механизмами, обеспечивающими охрану своей территории от вторжения чужаков, нехваткой мест, пригодных для устройства гнезд и нор, или же инфекционными заболеваниями.


 Зависимость силы влияния пресса хищников на популяцию жертв от плотности этой популяции была продемонстрирована (Messier, Crete, 1985) на примере волков и лосей в провинции Квебек (Канада). Выяснилось, что при численности лосей менее 0,2 особей/км2 пресс волков ничтожен. Волки не заселяют местности со столь низкой плотностью лосей или же полагаются на другие виды жертв. Когда плотность лосей достигает 0,2—0,5 особей/км2, пресс волков сильно возрастает и начинает играть важную роль в регуляции численности популяции. При дальнейшем же росте плотности популяции лосей (выше 0,5 особей/км2) она, перестает контролироваться волками, видимо, из-за того, что во-первых, нарушается социально-этологическая структура популяции волков (т. е. сложившаяся система взаимоотношений между семьями и отдельными особями в пределах каждой семьи), а во-вторых, потребности хищников восполняются в значительной степени за счет больных, ослабленных и прочих «нестандартных» особей жертвы, которые в любом случае не вносят сколь либо существенного вклада в размножение популяции или обеспечение ее более высокой выживаемости[45].
Зависимость силы влияния пресса хищников на популяцию жертв от плотности этой популяции была продемонстрирована (Messier, Crete, 1985) на примере волков и лосей в провинции Квебек (Канада). Выяснилось, что при численности лосей менее 0,2 особей/км2 пресс волков ничтожен. Волки не заселяют местности со столь низкой плотностью лосей или же полагаются на другие виды жертв. Когда плотность лосей достигает 0,2—0,5 особей/км2, пресс волков сильно возрастает и начинает играть важную роль в регуляции численности популяции. При дальнейшем же росте плотности популяции лосей (выше 0,5 особей/км2) она, перестает контролироваться волками, видимо, из-за того, что во-первых, нарушается социально-этологическая структура популяции волков (т. е. сложившаяся система взаимоотношений между семьями и отдельными особями в пределах каждой семьи), а во-вторых, потребности хищников восполняются в значительной степени за счет больных, ослабленных и прочих «нестандартных» особей жертвы, которые в любом случае не вносят сколь либо существенного вклада в размножение популяции или обеспечение ее более высокой выживаемости[45].
|
|
|
Возрастание среднего потребления пищи при увеличении количества ее в окружающей среде, как правило, имеет своим следствием повышение удельной рождаемости и снижение удельной смертности потребителя (хищника в широком смысле слова). Обе эти величины постепенно выходят на плато: максимальная рождаемость не может превышать некоторого предела, накладываемого допустимой частотой продуцирования потомков (детенышей, яиц, семян, спор и т. д.), а минимальная смертность не может быть меньше некоторого уровня всегда существующей в популяции физиологической смертности. Описывающие эту зависимость кривые (подобные той, что изображены на рис. 44) можно сравнительно легко получить в эксперименте для мелких быстро размножающихся организмов, например коловраток, потребляющих одноклеточные планктонные водоросли, или же для самих планктонных водорослей, потребляющих ионы фосфора или азота. Точке пересечения кривых рождаемости и смертности (т. е. состоянию равенства рождаемости и смертности) соответствует некоторая пороговая концентрация пищи (R*), при которой популяция сохраняет постоянную численность. При меньшей концентрации пищи численность популяции снижается (поскольку b < d), а при большей — растет (поскольку b > d). Подобные кривые нередко используются при анализе конкуренции между разными видами потребителей, и мы вернемся еще к их рассмотрению в соответствующем разделе данной главы.
|
|
|
