 |


Внутренние (висцеральные) анализаторы
|
|
|
|
Внутренние анализаторы осуществляют анализ и синтез информации о состоянии внутренней среды организма и участвуют в регуляции работы внутренних органов. Можно выделить: 1) внутренний анализатор давления в кровеносных сосудах и давления (наполнения) во внутренних полых органах (периферическим отделом этого анализатора являются механорецепторы); 2) анализатор температуры; 3) анализатор химизма внутренней среды организма; 4) анализатор осмотического давления внутренней среды. Рецепторы этих анализаторов расположены в различных органах, сосудах, слизистых оболочках и ЦНС.
К механорецепторам относятся все рецепторы, для которых адекватными стимулами являются давление, а также растяжение, деформация стенок органов (сосуды, сердце, легкие, желудочно-кишечный тракт и другие внутренние полые органы). К хеморецепторам относят всю массу рецепторов, реагирующих на различные химические вещества: это рецепторы аортального и каротидного клубочков, рецепторы слизистых оболочек пищеварительного тракта и органов дыхания, рецепторы серозных оболочек, а также хеморецепторы головного мозга. Осморецепторы локализованы в аортальном и каротидном синусах, в других сосудах артериального русла, в интерстициальной ткани вблизи капилляров, в печени и других органах. Часть осморецепторов является механорецепторами, часть — хеморецепторами. Терморецепторы локализованы в слизистых оболочках пищеварительного тракта, органов дыхания, мочевого пузыря, серозных оболочках, в стенках артерий и вен, в каротидном синусе, а также в ядрах гипоталамуса.
Проводниковый отдел. От интерорецепторов возбуждение восновном проходит в одних стволах с волокнами вегетативной нервной системы. Первые нейроны находятся в соответствующих чувствительных ганглиях, вторые нейроны — в спинном или продолговатом мозге. Восходящие пути от них достигают заднемеди-альное ядро таламуса (третий нейрон) и затем поднимаются в кору больших полушарий (четвертый нейрон).
|
|
|
Корковый отдел локализуется в зонах Ci и С2 соматосенсорной области коры и в орбитальной области коры большого мозга.
Восприятие некоторых интероцептивных стимулов может сопровождаться возникновением четких, локализованных ощущений, например при растяжении стенок мочевого пузыря или прямой кишки. Но висцеральная импульсация (от интерорецепторов сердца, сосудов, печени, почек и др.) может и не вызывать ясно осознаваемых ощущений. Обусловлено это тем, что такие ощущения возникают в результате раздражения различных рецепторов, входящих в ту или иную систему органов. В любом случае изменения внутренних органов оказывают значительное влияние на эмоциональное состояние и характер поведения человека.
Кожные анализаторы
Температурный анализатор
Температурный кожный анализатор обеспечивает информацию о температуре внешней среды и формирование температурных ощущений, что имеет большое значение для осуществления процессов терморегуляции и поведенческих приспособительных реакций. Как и тактильный, он относится к соматосенсорному анализатору.
Периферический отдел представлен двумя видами рецепторов: одни реагируют на холодовые стимулы, другие — на тепловые. Тепловые рецепторы — это тельца Руффини, а холодовые — колбы Краузе. Рецепторы холода расположены в эпидермисе и непосредственно под ним, а рецепторы тепла — преимущественно в нижнем и верхнем слоях собственно кожи и слизистой оболочки.
Проводниковый отдел. От рецепторов холода отходят миелинизированные волокна типа А, а от рецепторов тепла — немиелинизированные волокна типа С, поэтому информация от холодовых рецепторов распространяется с большей скоростью, чем от тепловых. Первый нейрон локализуется в спинальных ганглиях.
|
|
|
Клетки задних рогов спинного мозга представляют второй нейрон. Нервные волокна, отходящие от вторых нейронов температурного анализатора, переходят через переднюю комиссуру на противоположную сторону в боковые столбы и в составе латерального спинно-таламического тракта доходят до зрительного бугра, где находится третий нейрон. Отсюда возбуждение поступает в кору полушарий большого мозга.
Центральный отдел температурного анализатора локализуется в области задней центральной извилины коры большого мозга.
Восприятие температурных раздражителей. Существует зона температуры кожи, в пределах которой в результате адаптации к температуре внешней среды происходит полное исчезновение температурных ощущений. Эта зона получила название зоны комфорта, или нейтральной зоны. Изменения температуры кожи и отклонения от зоны комфорта происходят под влиянием факторов внешней и внутренней сред организма и сопровождаются возникновением ощущения тепла или холода. Интенсивность этих ощущений зависит от величины отклонения от диапазона зоны комфорта. Если температура кожи не меняется и какое-то время остается постоянной, то реакция терморецепторов в этих случаях обозначается как статическая. Уровень статической реакции зависит от длительности температурного раздражения и величины отклонения от диапазона зоны комфорта. При длительном воздействии температурных факторов внешней среды и малых отклонениях температуры кожи возможно развитие медленной частичной адаптации с сохранением низкого уровня статической реакции терморецепторов. При значительном изменении температуры внешней среды и больших отклонениях от зоны комфорта, когда развитие адаптации уменьшается, проявляется высокий уровень статической реакции терморецепторов.
Различают также динамические реакции терморецепторов, при которых формируются температурные ощущения, связанные с изменениями температуры кожи. Динамические реакции терморецепторов определяются тремя параметрами: исходной температурой и скоростью изменения температуры внешней среды, а также величиной поверхности кожи, на которую действует температурный фактор. Исходная температура кожи определяет уровень возбудимости терморецепторов: чем ниже температура кожи, тем выше возбудимость Холодовых и ниже — тепловых рецепторов и наоборот. При большой скорости изменения температуры внешней среды происходят быстрые изменения возбудимости терморецепторов кожи. При малой скорости изменения температуры среды возбудимость рецепторов изменяется медленно и может наблюдаться явление аккомодации, т.е. приспособление к воздействию медленно нарастающего температурного фактора, проявляющегося в снижении возбудимости терморецепторов кожи. Интенсивность температурных ощущений находится в прямо пропорциональной зависимости от величины поверхности кожи, на которую воздействует температурный стимул: чем больше площадь воздействия температурного фактора, тем температурные ощущения сильнее, и наоборот, если маленькие участки кожи подвергаются воздействию температуры, ощущения понижены. Это явление объясняют наличием пространственной суммации на разных уровнях проводникового отдела температурного анализатора, что оказывает влияние на формирование температурных ощущений.
|
|
|
Данное объяснение подтверждается опытом с двусторонней стимуляцией. Так, например, при одновременном температурном воздействии на тыльную поверхность обеих рук температурные ощущения будут выше, чем при обогревании или охлаждении одной руки.
Иногда наблюдаются парадоксальные ощущения холода при воздействии высоких температур. Это можно объяснить тем, что холодовые рецепторы располагаются ближе к поверхности кожи (на глубине 0,17 мм), чем тепловые, расположенные на глубине 0,3 – 0,6 мм, поэтому холодовые рецепторы возбуждаются быстрее. В то же время считают, что причина этого явления, возможно, лежит в том, что холодовые рецепторы, в норме «молчащие» при температуре выше +40° С, вдруг возбуждаются на короткое время, если на них быстро подействовать температурой выше +45° С.
|
|
|
Тактильный анализатор
Тактильный анализатор является частью кожного анализатора. Он обеспечивает ощущения прикосновения, давления, вибрации и щекотки.
Периферический отдел представлен различными рецепторными образованиями, раздражение которых приводит к формированию специфических ощущений. На поверхности кожи, лишенной волос, а также на слизистых оболочках на прикосновение реагируют специальные рецепторные клетки (тельца Мейснера), расположенные в сосочковом слое кожи. На коже, покрытой волосами, на прикосновение реагируют рецепторы волосяного фолликула, обладающие умеренной адаптацией.
На давление реагируют рецепторные образования (диски Меркеля), расположенные небольшими группами в глубоких слоях кожи и слизистых оболочек. Это медленно адаптирующиеся рецепторы. Адекватным стимулом для них служит прогибание эпидермиса при действии механического стимула на кожу.
Вибрацию воспринимают тельца Пачини, располагающиеся как в слизистой, так и на не покрытых волосами частях кожи, в жировой ткани подкожных слоев, а также в суставных сумках, сухожилиях. Эти рецепторы представлены нервными терминалями, заключенными в слоистые оболочки из соединительной ткани. Тельца Пачини обладают очень быстрой адаптацией и реагируют на ускорение при смещении кожи в результате действия механических стимулов, одновременно вовлекаются в реакцию несколько телец Пачини.
Щекотание воспринимают свободно лежащие, неинкапсулированные нервные окончания, расположенные в поверхностных слоях кожи. Для данного вида рецепторов характерна низкая специфичность реакции на стимулы разной интенсивности. С активацией этой группы рецепторов связывают ощущение щекотки, что и дало название самим рецепторам — рецепторы щекотки.
По функциональным особенностям тактильные рецепторы подразделяются на фазные и статические. Фазные тактильные рецепторы возбуждаются при динамическом раздражении. Они обладают высокой чувствительностью, коротким латентным периодом, быстро адаптируются. Статические тактильные рецепторы возбуждаются в основном от статического раздражения. Они менее чувствительны, чем фазные, с более длительным латентным периодом, медленно адаптируются.
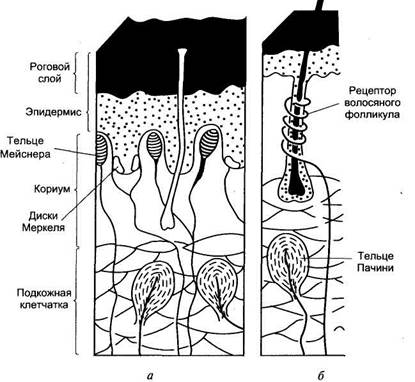
Рисунок: схема строения и положения механорецепторов в коже на не покрытых волосами (а) и волосистых (б) участках кожи
Проводниковый отдел. От большинства механорецепторов в спинной мозг информация поступает в центральную нервную систему по А-волокнам и лишь от рецепторов щекотки — по С-волокнам. Первый нейрон находится в спинальных ганглиях. В заднем роге спинного мозга происходит первое переключение на интернейроны (второй нейрон), от них восходящий путь в составе заднего столба достигает ядер заднего столба в продолговатом мозге (третий нейрон), где происходит второе переключение, далее через медиальную петлю путь следует к вентро-базальным ядрам зрительного бугра (четвертый нейрон), центральные отростки нейронов зрительного бугра идут в кору больших полушарий.
|
|
|
Центральный отдел тактильного анализатора локализуется в I и II зонах соматосенсорной области коры большого мозга (задняя центральная извилина).
Исследования уровня тактильной чувствительности можно проводить с помощью «волосков Фрея», а пространственных порогов, которые характеризуют плотность распределения тактильных рецепторов, — «циркулем Вебера».
Болевой анализатор
Болевой анализатор осуществляет формирование болевых, ощущений (боли), которые возникают при воздействии повреждающих факторов.
Боль — это «сенсорная модальность» подобно слуху, вкусу, зрению и пр., она выполняет сигнальную функцию, которая заключается в информации о нарушении таких жизненно важных констант организма, как целостность покровных оболочек и определенный уровень окислительных процессов в тканях, обеспечивающих их нормальную жизнедеятельность.
В то же время боль можно рассматривать как психофизиологическое состояние, сопровождаемое изменениями деятельности различных органов и систем, а также возникновением эмоций и мотиваций.
Структурно-функциональная характеристика
Периферический отдел анализатора представлен рецепторами боли, которые по предложению Ч. Шеррингтона называют ноцицепторами (от лат. разрушать). Это высокопороговые рецепторы, реагирующие на разрушающие воздействия. По механизму возбуждения ноцицепторы делят на механоноцицепторы и хемоноцицепторы.
Механоноцицепторы расположены преимущественно в коже, фасциях, сухожилиях, суставных сумках и слизистых оболочках пищеварительного тракта. Это свободные нервные окончания миелинизированных волокон типа А-дельта со скоростью проведения возбуждения 4 — 30 м/с. Они реагируют на действие агента, вызывающего деформацию и повреждение мембраны рецептора присжатии или растяжении тканей. Для большинства этих рецепторов характерна быстрая адаптация.
Хемоноцицепторы расположены также на коже и в слизистых оболочках, но превалируют во внутренних органах, где локализуются в стенках мелких артерий. Они представлены свободными нервными окончаниями немиелинизированных волокон типа С со скоростью проведения возбуждения 0,4 — 2 м/с. Специфическими раздражителями для этих рецепторов являются химические вещества (алгогены), но только те, которые отнимают кислород у тканей, нарушают процессы окисления.
Выделяют три типа алгогенов, каждый из которых имеет собственный механизм активации хемоноцицепторов. Тканевые алгогены (серотонин, гистамин, ацетилхолин и др.) образуются при разрушении тучных клеток соединительной ткани и, попадая в интерстициальную жидкость, непосредственно активируют свободные нервные окончания. Плазменные алгогены (брадикинин, каллидин и простагландины), выполняя роль модуляторов, повышают чувствительность хемоноцицепторов к ноцигенным факторам. Тахикинины выделяются при повреждающих воздействиях из окончаний нервов (к ним относится вещество полипептид), они воздействуют местно на мембранные рецепторы того же нервного окончания.
Представление о наличии специфического сложноорганизован-ного периферического отдела болевого анализатора основано на теории специфичности (М.Фрей, 1895). Согласно этой теории, ощущение боли может возникать только при раздражении определенных рецепторов (ноцицепторов) и специальных путей проведения возбуждения в соответствующие нервные центры. Однако боль может возникать также при очень сильных, в том числе повреждающих воздействиях и на другие рецепторы, например терморецепторы. Такой взгляд сформировался на основе теории интенсивности (А. Гольдштейдер, 1894) — неспецифической теории боли. Согласно этой теории, ощущение боли формируется в результате суммации в нервных центрах возбуждений, возникающих при раздражении рецепторов различных модальностей.
В настоящее время считают, что боль может возникать как при повреждающих воздействиях на специальные рецепторы — ноцицепторы, так и при интенсивном воздействии на рецепторы различной модальности. Развивается также представление, согласно которому под термином «ноцицептор» подразумевается не только сам рецепторный аппарат, но и связанная с ним периферическая часть афферентного волокна. Это образование получило название «сенсорной болевой единицы». Предполагается, что сенсорная болевая единица имеет два возбудимых участка. Один из них (претерминальная часть дендрита) возбуждается только повреждающими стимулами. Другой (сама терминаль) может активизироваться воздействиями, не несущими ноцицептивной информации (субноцицептивными воздействиями).
Проводниковый отдел. Проведение болевого возбуждения от рецепторов осуществляется по дендритам первого нейрона, расположенного в чувствительных ганглиях соответствующих нервов, иннервирующих определенные участки организма. Аксоны этих нейронов поступают в спинной мозг к вставочным нейронам заднего рога (второй нейрон). Далее проведение возбуждения в центральной нервной системе осуществляется двумя путями, специфическим (лемнисковым) и неспецифическим (экстралемнисковым) Специфический путь начинается от вставочных нейронов спинного мозга, аксоны которых в составе спиноталамического тракта поступают к специфическим ядрам таламуса (в частности, в вентробазальное ядро), которые представляют третьи нейроны. Отростки этих нейронов достигают коры.
Неспецифический путь начинается также от вставочного нейрона спинного мозга и по коллатералям идет к различным структурам мозга. В зависимости от места окончания выделяют три основных тракта — неоспиноталамический, спиноретикулярный, спиномезенцефалический.
Последние два тракта объединяются в спиноталамический. Возбуждение по этим трактам поступает в неспецифические ядра таламуса и оттуда во все отделы, коры больших полушарий.
Корковый отдел. Специфический путь заканчивается в соматосенсорной области коры большого мозга. Согласно современным представлениям выделяют две соматосенсорные зоны. Первичная проекционная зона находится в области заднецентральной извилины. Здесь происходит анализ ноцицептивных воздействий, формирование ощущения острой, точно локализованной боли. Кроме того, за счет тесных связей с моторной зоной коры осуществляются моторные акты при воздействии повреждающих стимулов. Вторичная проекционная зона, которая находится в глубине сильвиевой борозды, участвует в процессах осознания и выработке программы поведения при болевом воздействии.
Неспецифический путь распространяется на все области коры. Значительную роль в формировании болевой чувствительности играет орбитофронтальная область коры, которая участвует в организации эмоционального и вегетативного компонентов боли.
Компоненты реакции организма на боль. В реакцию организма на боль вовлекаются практически все структуры головного мозга, поскольку по коллатералям проводникового отдела болевого анализатора возбуждение распространяется на ретикулярную формацию, лимбическую систему мозга, гипоталамус и двигательные ядра. В связи с этим в реакции организма на боль выделяют несколько компонентов. Двигательный компонент проявляется при включении мотонейронов и обнаруживается в виде отдельных двигательных рефлексов, реакций вздрагивания и настороженности, а также защитного поведения, направленного на устранение действия вредоносного фактора. Вегетативный компонент обуслорлен включением в системную болевую реакцию гипоталамуса — высшего вегетативного центра. Этот компонент проявляется в изменении вегетативных функций, необходимых для обеспечения защитной реакции организма. В зависимости от индивидуальных особенностей организма, его вегетативного статуса могут наблюдаться реакции с разнонаправленными изменениями величин артериального давления, частоты сердечных сокращений, дыхания, перестройки обмена веществ и т.д. Эмоциональный компонент проявляется в формировании отрицательной эмоциональной реакции, что обусловлено включением в процесс возбуждения эмоциогенных зон мозга. В зависимости от индивидуально-генетических особенностей организма, и в частности вегетативного тонуса, отрицательная эмоция формирует различные поведенческие реакции, например бегство или нападение. В организации защитных поведенческих реакций ведущая роль отводится структурам лобной и теменной областей коры полушарий большого мозга.
Каждый компонент болевой реакции может быть использован для оценки специфичности болевого ощущения при медицинских и биологических исследованиях.
|
|
|
