 |


1.4 Токи мышц и нервов.
|
|
|
|
Самые наглядные электрические явления наблюдаются в мышцах и нервах, токи которых могут быть отведены к гальванометру и произвести в этом последнем более или менее значительное отклонение магнитной стрелки. Уже Маттеуччи наблюдал электрические токи в мышцах, но только Дю-Буа-Реймон определил условия их происхождения и формулировал законы, по которым эти токи распространяются. Если поперечный разрез и продольную поверхность любой мышцы лягушки (лучше всего мышцы с параллельно расположенными волокнами, напр., толстой трехглавой мышцы бедра) соединить посредством неполяризуюшихся электродов с гальванометром, то наблюдается в этом последнем более или менее значительное отклонение стрелки, которое указывает на несомненное существование тока в исследуемой мышце. Более того, из направления, в котором отклоняется стрелка, можно заключить, что наблюдаемый ток направляется в мышце от поперечного разреза к её продольной поверхности, следовательно, в гальванометрической цепи, т. е. по проводникам, соединяющим мышцу с гальванометром, ток идет от продольной поверхности мышцы к её поперечному разрезу. Этот ток носит название поперечно-продольного тока мышцы и указывает на то, что поперечный разрез мышцы относится отрицательно к её продольной поверхности, которая наэлектризована положительно. Самый сильный ток идет по направлению от середины поперечного разреза мышцы к её экватору, т. е. к середине её продольной поверхности. В этих двух точках находится максимум электрического потенциала, который убывает по мере удаления от этих точек. Таким образом, более слабые токи получаются от соединения двух точек продольной поверхности, находящихся в неодинаковом расстоянии от экватора или же от соединения двух точек поперечного сечения, неодинаково отстоящих от центра. Две точки, находящиеся на равных расстояниях от середины продольной поверхности мышцы или от центра её поперечного разреза, не представляют разницы электрического потенциала, следовательно, не обнаруживают никакого тока. Совокупность всех этих явлений, уже отчасти известных Маттеучи, составляют закон мышечных токов, который был впервые формулирован Дюбуа-Реймоном. Следующая схема показывает силу и направление токов в мышце, снабженной двумя поперечными разрезами и представляющей вид цилиндрической призмы.
|
|
|
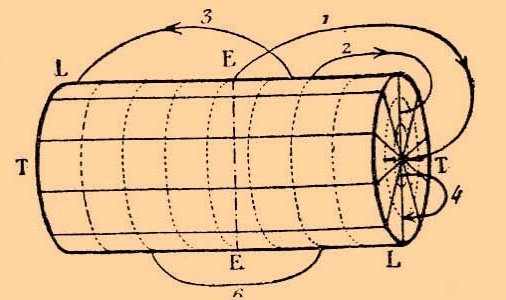
Рис 2. - Распределение электрических потенциалов в правильном цилиндре, изображающем мышцу, снабженную двумя поперечными разрезами: ЕЕ — экватор; ТТ — поперечные сечения; L — продольная поверхность; 1, 2, 3, 4 — мышечные токи, направление которых обозначено стрелками; 6 — отсутствие тока между двумя точками продольной поверхности, одинаково отстоящими от экватора.
Если вместо поперечных разрезов, перпендикулярных к продольной оси мышцы, провести через мышцу два косые поперечные разреза, то вместо цилиндра получится ромб, в котором распределение токов совсем другое, чем в нормальном мышечном цилиндре. Все точки поперечного разреза или продольной поверхности ромба, лежащие ближе к тупому углу, относятся электроположительно ко всем точкам, лежащим ближе к острому углу. Получаемые при таком распределении электрических потенциалов токи называются токами наклонения; они обыкновенно сильнее токов правильного мышечного цилиндра. Некоторые нормальные мышцы имеют тоже ромбоидальную форму в силу особенного расположения своих волокон. Икроножная мышца лягушки представляет даже двойной мышечный ромб; её сухожильная часть, составляющая естественный поперечный разрез мышцы, является электроположительной в отношении к продольной поверхности. Своеобразное направление токов в такой мышце, резко отличающееся от направления токов в нормальной мышце, составляет, по Дюбуа-Реймону, явление парэлектрономии мышц. Достаточно сделать искусственный поперечный разрез, чтобы получить правильное распределение электрического потенциала. Выше описанная законность в распределении электрических токов свойственна не только целой мышце, но наблюдается также в её отдельных кусках. Этот факт указывает на то, что мышечный ток есть явление, присущее каждому отдельному мышечному волокну. Электродвигательная сила мышечного тока равна приблизительно 0, 03—0, 08 вольта и тесно связана с жизненными свойствами мышцы. Она ослабевает в умирающей мышце и вполне исчезает при наступлении трупного окоченения. В нормальной свежей мышце сила мышечного тока достаточна для того, чтобы раздражать нерв и вызвать сокращение в нервно-мышечном аппарате. Это явление было описано уже давно самим Гальвани под названием: " сокращений без металлов".
|
|
|
Токи в нервах во время покоя. Токи в нервах, открытые впервые Дюбуа-Реймоном (1844), распределяются таким же образом, как и в мышце. Поперечный разрез нерва относится электроотрицательно к его естественной продольной поверхности. Середина продольной поверхности (экватор) нерва и центр его поперечного сечения представляют максимум электрического потенциала, который ослабевает по мере удаления от этих точек.
Все токи, наблюдаемые в нерве, сводятся к следующим трем типам:
1) ток поперечно-продольный, идущий от поперечного разреза нерва к его продольной поверхности;
2) ток продольной поверхности получается между двумя точками этой поверхности, лежащими в неодинаковом расстоянии от экватора, и
3) осевой ток, направляющийся от одного поперечного разреза к другому.
Каждый из двух поперечных разрезов нерва относится, как было оказано выше, электроотрицательно к продольной поверхности, но оба поперечные разреза, сравниваемые между собой, не представляют одинакового электрического потенциала. В двигательных, т. е. центробежных, нервах поперечный разрез центрального конца является менее электроотрицательным, значит, положительным по отношению к поперечному разрезу периферического конца. Наоборот, в чувствительных, т. е. центростремительных, нервах периферический поперечный разрез относится положительно к центральному поперечному сечению. Из этого следует, что осевой ток имеет восходящее направление в центробежных (двигательных) нервах и нисходящее направление в центростремительных (чувствительных) нервах, значит, направление осевого тока противоположно направлению физиологической функции нерва. Этот закон осевого тока имеет большой теоретический интерес, так как он прямо указывает на функциональную связь животного электричества с физиологической деятельностью нерва. Направление осевого тока в нерве составляет тоже весьма наглядную и даже единственную разницу между центростремительными и центробежными нервами. Электродвигательная сила нервного тока равна 0, 005—0, 03 вольта; она зависит от жизненности нерва и от его толщины. Хотя сила нервного тока сравнительно весьма незначительна, но она достаточна для того, чтобы произвести раздражение нерва в другом нервно-мышечном препарате. Замкнув цепь поперечно-продольного тока нерва, можно вызвать так же, как и в мышце, самораздражение данного нерва и, следовательно, сокращение мышцы, иннервируемой этим нервом. Если произвести такое же побочное замыкание поперечно-продольного тока в электрическом нерве ската, то можно получить во время замыкания и размыкания тока довольно сильный разряд электрического органа.
|
|
|
Токи действия мышц и нервов. Все вышесказанное относится к покоящимся мышцам и нервам. Но и во время действия мышца и нерв обнаруживают токи, впервые описанные Дюбуа-Реймона и названные им отрицательным колебанием мышечного и нервного тока. По Дюбуа-Реймону, ток, происходящий во вредя действия мышц и нервов, влечет за собой уменьшение их собственного тока, между тем как по Херману, на эти токи надо смотреть прямо как на токи действия, связанные с процессом возбуждения в мышце и нерве. Если отвести к гальванометру собственный поперечно-продольный ток мышцы или нерва и раздражать эти последние токами, достаточно сильными для того, чтобы произошло мышечное сокращение, то легко можно убедиться в том, что при переходе мышцы или нерва в состояние действия, магнитная стрелка двигается обратно по направлению к нулю. Если она до раздражения остановилась, напр., на 60-м делении шкалы, то в момент мышечного сокращения она может, двигаясь в обратном направлении, остановиться на 30-м или даже на 20-м делении шкалы. Это явление несомненно указывает на то, что собственный ток покоящейся мышцы или нерва значительно ослабел во время их действия и что это ослабление есть результат развития тока обратного направления, составляющего именно отрицательное колебание покойного тока. Иногда отрицательное колебание может быть столь сильным, что совершенно уничтожает покойный ток; магнитная стрелка возвращается тогда к нулю. Продолжительность отрицательного колебания равна приблизительно 1/300 секунды. Оно распространяется со скоростью 3 метров в секунду. Отрицательное колебание может произвести в другом нервно-мышечном препарате раздражение и сокращение мышцы. Лягушечья лапка, расположенная на бьющемся сердце кролика, раздражается и сокращается от тока действия каждого удара сердца. Во время тетанического, т. е. длительного сокращения мышцы, происходящего от действия известного количества прерывистых раздражений, каждое одиночное раздражение сопровождается соответственным отрицательным колебанием, которое можно легко проследить гальванометрически или электрометрически, или же с помощью телефона. Сила отрицательного колебания в известных границах пропорциональна силе покойного тока. Отрицательное колебание мышечного тока находится в тесной связи с силой сокращения мышцы: в нерве же отрицательное колебание нервного тока служит лучшим и даже единственным средством для обнаруживания деятельности нерва, если нервное возбуждение не сопровождается мышечным сокращением.
|
|
|
Электрические токи в других тканях и органах. — Уже Дюбуа-Реймон доказал существование электрических токов в коже, наружная поверхность которой является электроположительной в отношении к внутренней поверхности; таким образом ток кожи направляется снаружи внутрь. Подобные электродвигательные свойства представляют тоже разные слизистые оболочки, снабженные железами и даже кожа рыб, лишенная желез. Электрические тока желез обнаруживаются не только при покое, но и во время их действия (секреторные токи). Выделение слюнных желез сопровождается током действия. Такой же ток наблюдается и во время секреторной деятельности слизистой оболочки желудка.
|
|
|
Нервные центры также обнаруживают различие электрических потенциалов, хотя токи в спинномозговой оси не представляют того правильного и строго законного распределения, как в мышце и нерве. Это связано, конечно, с весьма сложным строением центральной нервной системы. В продолговатом и спинном мозгу токи покоя и токи действия были впервые наблюдаемы Сеченовым. Такие же токи были определены другими экспериментаторами и в головном мозгу. Токи действия в этом органе происходят не только от непосредственного возбуждения самого вещества мозга, но и от раздражения любого центростремительного нерва. Передняя половина мозга (лобные доли) относится электроположительно к его задней половине (затылочные доли); но вообще направление токов в головном мозгу весьма разнообразно и не может быть подведено под одну общую схему. Тем не менее законы происхождения и paпределения электрических токов, сопровождающих деятельность головного мозга, составляют несомненно одну из самых важных задач психо-физиологии. К сожалению, вопрос этот еще недостаточно разработан и известные в этом отношении факты не допускают никаких общих заключений. В органах чувств наблюдаются также электродвигательные явления, которые особенно хорошо изучены в глазу. Юыло доказоно, что зрительный нерв у рыб относится электроотрицательно к передней части (к роговице) глазного яблока. Электрические явления в глазу могут быть вызваны с помощью адекватного, т. е. светового раздражения, причем желтый свет действует сильнее всего в электрогенном смысле, другие цвета действуют слабее. Токи действия находятся, по всей вероятности, в связи с зрительным пурпуром сетчатки; они значительно ослабевают в сетчатке с обесцвеченным пурпуром. Фото-электрические явления могут быть вызваны не только вследствие прямого действия света на исследуемый глаз, но и путем рефлекса, вследствие светового раздражения другого глаза.
|
|
|
