 |


Грань между живыми и неживыми природными системами
|
|
|
|
ОБЩИЕ ПРОБЛЕМЫ
ЭКОЛОГИЧЕСКОЙ БИОЛОГИИ
3.1. ЧТО НОВОЕ ПРИВНЕСЛИ МИКРООРГАНИЗМЫ В ТЕОРИЮ ЭВОЛЮЦИИ
Cслияние генетики с неодарвинизмом в 30—40-е годы прошлого века стараниями Добжанского, Майра и Симпсона привело к созданию синтетической теории эволюции (СТЗ). Было дано определение видов как «группы скрещивающихся естественных популяций, изолированные от таких же популяций репродуктивными барьерами» [Майр, 1974]. Недостаточные знания мира микробов не позволили в СТЗ большое внимание уделять микроорганизмам. Этим страдают и более поздние работы по эволюции, так как не всегда однозначно можно было трактовать исследования по мутациям бактерий, в которых прослеживались признаки ламаркизма.
Большой вздох облегчения пронесся, когда сначала Дельбрюк и Лурия в своем «флуктуационном тесте» на модели бактерия—фаг показали, что мутации в бактериях случайные и ненаправленные (цит. по [Ратнер, 1972]), а затем супруги Ледерберги продемонстрировали это наглядно, используя «метод отпечатков». Было даже высказано пафосное выражение, что в природе смыт последний «островок ламаркизма».
В настоящее время появилась устойчивая тенденция (если не мода) к пересмотру всех классических позиций эволюционной теории. Основной пищей для этого пересмотра явились экспериментальные данные, полученные в мире микроорганизмов. Какие же особые свойства присуши микроорганизмам, что обеспечивает существенные отличия от мира микроорганизмов? Чтобы не пересказывать весь курс микробиологии, мы выделим лишь свойства, важные, по мнению авторов, дня эволюционной теории.
Первое из них очевидно — это размер. Различие микро- и макроорганизмов условно. Объект микробиологии микроорганизмы, по определению, это не видимые невооруженным глазом живые существа. Действительно, линейные размеры прокариотических клеток редко превышают 20 мкм. Хотя есть микробы размера песчинок, например, Epulopiscium fishelsoni и особенно Thiomargarita namibiensis, достигающие 0,75 мм. В нашем случае важно, что в предельно ограниченном пространстве могут в тесном контакте находиться огромное количество микроорганизмов. В одной чашке Петри на агаризованной питательной среде за сутки вырастают микробы, по численности превышающие население Земли. Но чашка Петри — это искусственно созданные условия. В действительности, естественных экологических ниш со своими специфическими условиями невообразимо много.
|
|
|
И при значительном влиянии именно этих меняющихся по времени условий происходят эволюционные изменения микроорганизмов, приводящие к их многообразию. Из чего возникло это разнообразие, как и по каким законам оно возникло, почему именно такое и что будет дальше? Приземлим эти вопросы до ранга нашей планеты, хотя эксплуатация феномена «панспермия» может дать отсрочку нашим умозаключениям до поступления проясняющих знаний, перенося происхождение жизни за пределы Земли. Следует отметить, что это не покрывает нашу леность, а обусловлено рядом замечательных наблюдений в сохранившихся и поступающих фрагментах внеземного материала [Розанов, 1999]. В некоторых метеоритах, особенно в углистых хондритах типа Murchison, встречались микроструктуры, напоминающие микроколонии и отдельные клетки, причем часть из них имела морфологию, схожую с современными цианобактериями Enthophysalis granulosa, т.е. в достаточной степени являются сложной и специализированной структурой.
Вернемся к нашей планете, исходя из того, что ее возраст оценивается приблизительно в 4,8 млрд лет. Вначале был огненный остывающий шар, представить в котором развитие сложных структур трудно. Практически не вызывает сомнений тезис, что первыми и единственными обитателями Земли были микроорганизмы, способные поддерживать свою структуру в этих термах, наиболее близкие к экстремофилам, до сих пор существующим в особых экологических нишах нашей планеты. Этот период длился достаточно долго и привел к образованию атмосферы. Далее ─ симбиотическое происхождение эукариот, разнонаправленная эволюция жизненных форм, выход на сушу, освоение новых ниш. Все это наши умозаключения, так как микроорганизмы в силу своеобразия строения не имеют палеонтологической летописи.
|
|
|
Две других удивительных особенности микроорганизмов: «потенциальное бессмертие» и высокие темпы размножения при благоприятных условиях. Бактерии размножаются в основном бинарным делением, а это приводит к тому, что после всего цикла образуются с максимально возможной, контролируемой всей клеткой точностью, две копии, два клона, две дочерние клетки. Нет материнской, далее отмирающей клетки, а две равноценные дочерние. То есть бактерии, живущие в настоящий момент времени, — это именно те древние клетки, появившиеся на заре жизни и несущие в своем геноме все удары прошлого.
Огромные темпы размножения позволяют увеличить долю полезных случайно и не случайно появившихся вариантов за незначительное время. Минимальное время удвоения быстрорастущих клеток оценивается в 15 мин., а медленнорастущих — сутки (хотя все зависит от условий). То есть для быстрорастущих клеток за рабочую неделю пройдет период по смене поколений, эквивалентный более 10 тыс. лет для человека. Микробам не требуются значительные временные промежутки, исчисляемые годами, чтобы узаконить полезные для данной экологической ниши признаки. Причем появление новых генных систем напрямую зависит от той экологической обстановки, в которой оказался микроорганизм, а точнее, случайный набор популяций различных микроорганизмов. Наличие стабильных условий приводит к образованию стабильных трофических цепей, к значительному однообразию и лишь небольшим вариациям. Изменение же условий в стабильных нишах неизбежно приводит к изменению количественного или качественного состава участников трофических цепей. Это еще одно из важных отличий эволюции микроорганизмов: значительное, первостепенное влияние условий среды на направленность эволюционных изменений. Причем изменение условий не приводит к элиминации части микроорганизмов, не подходящих для размножения в изменившихся условиях. Особое свойство (более подробно описанное ниже) микробов переходить в длительное анабиотическое состояние позволяет сохранять эту часть, казалось бы, ненужных микроорганизмов, а с ними их генный потенциал.
|
|
|
Возможность секвенирования ДНК открыла перспективы создания филогене-тических систем. Это по своей сути редукционистское понятие, когда все явления биологии объясняют в терминах химии и физики. При этом предполагалась изоморфность филогении организмов, филогении молекул и всей таксономической системы. Конечно, фиксировались мутации, которые, как всем верилось, случайные и неадаптивные. Так как, несмотря на С-парадокс, геном микроорганизмов все же меньше, чем у эукариотических организмов, больше данных было получено и обработано по микроорганизмам, и, в конце концов, метрономом эволюции была выбрана последовательность гена малой рибосомальной РНК, 1 6S рРНК — для прокариот и 18S рРНК для эукариот [Woese, 1987]. Она (последовательность гена) имеет давнее историческое прошлое, присуща всем организмам, консервативна и насчитывает полторы тысячи нуклеотидов, т. е., перефразируя Н. Гоголя, нельзя сказать, чтобы коротка, однако ж и не так чтобы слишком длинна. Данные по последовательностям накапливались стремительно. Эволюционные построения древ стали делом техники. Прошел и по-прежнему идет процесс массовой ресистематизации, главным результатом которого стало вычленение целого царства живого, царства Архебактерий. Основным критерием стала доля схожести, или частоты замен, в последовательности малой рибосомальной РНК. Компьютерные программы стали определять принадлежность организма к тому или другому роду и виду. Сам организм с его другими физиологические свойствами стал не важен, вырван из среды обитания. Поэтому неудивительно, что вместо традиционных названий крупных таксонов появились такие: альфа-, бета-, гамма- и т. д. протеобактерии. Хватит ли в конечном счете всего греческого алфаита?
|
|
|
Первый удар по, казалось бы, стройной системе был нанесен совершенно неожиданно. Появился ряд работ, доказывающих адаптивность мутаций у микроорганизмов, сначала у бактерий, а затем и у дрожжей. Работа Каэрнса опровергала одну из фундаментальных предпосылок большинства эволюционных построений [Саirnc еt аI., 1988]. Было показано, что мутации, происходящие в генах, не случайны, а зависят от условий окружающей среды, в которой эти микроорганизм оказались. Каэрнс с коллегами показали, что модель в классических опытах по ненаправленности мутаций была подобрана некорректно. Действие литического бактериофага Т1 на Escherichia coli было настолько быстро, что не позволяло реализовать адаптивные возможности бактерии в виде повышения частоты полезных мутаций. Более адекватной представлялась модель также со штаммом Е. coli, но имеющим делецию в гене бета-галактозидазы. Авторы продемонстрировали, что в специальных условиях, когда единственным источником углерода в питательной среде была лактоза, т. е. в норме клетки не могли размножаться, частота Iас+-фенотипа значительно возрастает. Более того, для нейтрального признака, не связанного с селекцией по лактозе, частота мутаций была даже ниже. То есть при создании селективных условий не только повышается частота полезных, но и снижается частота нейтральных мутаций. Организм ориентирован, направлен на преодоление неблагоприятных условий, даже в условиях не размножающихся клеток.
Заключение из перечисленных выше исследований неутешительно: теория нейтральности эволюции не применима (по крайней мере, не в каждом случае применима) к миру микроорганизмов. Это одно из основных отличий эволюции микроорганизмов от эволюции макроорганизмов. Не всегда, казалось бы, незыблемые законы можно напрямую переносить в мир, живущий и подчиняющийся другим правилам и законам.
Что еще необходимо отметить как особенность, так это другая сторона медали быстрых темпов размножения в благоприятных условиях. для микроорганизмов существуют документально доказанные периоды стагнации, анабиоза. Это время, в котором, по нашим представлениям, с микроорганизмом ничего не происходит, так как он не имеет возможностей для размножения. При этом сохраняется жизнеспособность и при попадании в благоприятные условия микроб просыпается из спячки и продолжает свое существование дальше. Удивителен не сам факт такого состояния, все помнят про медведя, а его продолжительность. Представлены теперь уже многочисленные факты возможности выхода из анабиоза после тысячелетий, миллионов, десятков и даже сотен миллионов лет консервации. Мы же в своих работах отмечали, что нельзя представлять состояние стадии переживания неблагоприятных условий как отсутствие метаболизма бактерий вообще. При анализе микроорганизмов, выделенных из мозга юкагирского мамонта, находящегося более 18 тыс. лет в законсервированном вечной мерзлотой состоянии, были обнаружены несколько морфотипов бактерий [Repin et al., 2007]. Они были как на 100 % схожие с современными, так и отличающиеся от них комбинаторным перемешиванием имеющихся в наличии признаков (табл. 3.1).
|
|
|
Представляется, однако, существенным, что если геномные перестройки микроорганизмов идут в таких специфических условиях, то, по крайней мере, их темпы замедлены. Несомненно, что в мире одновременно существуют микроорганизмы, возраст которых различается в миллионы лет. Для осмысления временных параметров напоминаем, что возраст человечества, беря отсчет с гоминид, оценивается в несколько миллионов лет. Это время, которое некоторые микроорганизм просто проспали.
При этом следует сказать, что оживление «древних» микроорганизмов показало, что существуют стабильные состояния, отточенные эволюцией. Эти «древние» бактерии имели набор признаков и последовательности гена 16S рРНК, полностью совпадающие с современными, будь то микроорганизмы из мозга мамонта или бациллы, выделенные из дрозофилы, законсервированной в янтаре [Саnо,Boruchi, 1995].
Настало время от порядка вернуться к комбинаторным перестановкам, «пространству логических возможностей» [Заварзин, 1974]. На заре эры всеобщего секвенирования предполагалось, что в мире микроорганизмов, конечно, происходит обмен генетической информацией между видами и даже родами. Однако обмен все же ограничен и описывается известными процессами, конъюгацией, трансформацией, трансдукцией и т. д. Опубликование последовательностей, в том числе и полных, закрыло вопрос о значительной, преобладающей роли обменов ДНК между микроорганизмами, по-видимому, навсегда (см. табл. 3.1). Теперь мы видим,
Таблица 3.1
Горизонтальный перенос генов у архей и бактерий
| Вид | Число генов в геноме | Перенесенные гены | |
| количество | % в геноме | ||
| Археи Methanococcus jannaschii Pyrococcus horikoshii Aeropyrum pernix Патогенные Mycoplasma pneumoniae Chlamydia trachomatis Rickettsia prowazekii Treponema pallidum Haemophilus influenzae Helicobacter pylori Mycobacterium tuberculosis Свободноживущие бактерии Thermotoga maritime Escherichia coli Pseudomonas aeruginosa Bacillus subtilis | 5,0 7,6 14,0 5,9 4,3 3,6 8,3 6,2 6,4 5,0 11,6 9,6 10,1 14,8 |
что, по крайней мере для микроорганизмов, нет принципиальных барьеров (хотя, например, РМ-системы имеются) для обмена генетической информацией, включая обмены между царствами, и эти обмены существенны даже в процентном отношении ко всему конкретному геному. Наиболее подходящей схемой, по нашему представлению, является положение о «пространстве логических возможностей», гениально предложенное академиком Г. А. Заварзиным в начале восьмидесятых годов прошлого века [Заварзин, 1974]. В мире микроорганизмов потенциально могут существовать комбинации генов, не противоречащих, совместимых между собой и всей системой. Тогда эволюционную схему можно будет представить как подсолнечник, где некоторые семечки выклеваны воронами, или рыболовную сеть с некоторыми рваными ячейками, в которую не может попасть рыба [Repin, Vlassov, 2007]. В качестве фактов, подтверждающих данную теорию, можно показать наши последние работы, представленные в этой монографии. Существуют ряд незыблемых положений, например: споры могут образовываться только грамположительными бактериями, и то не всеми. Учитывая, что за спорообразование ответствен не один десяток генов, трудно предположить, что получение всего кластера — быстрый процесс. Однако, исследуя термофилы Забайкалья и, далее, Камчатки, нам удалось выделить такие грамотрицательньге спорообразующие бактерии. функционирование комбинации генов, обусловливающих структуру клеточной стенки и спорообразование, оказалось совместимым друг с другом [Аndreeva еt аI.,2007]. Для систематики прокариотических микроорганизмов это может иметь трагические последствия: огромное, не поддающееся на данный момент времени анализу разнообразие. Каждому штамму будет присвоен штрих-код; Однако, учитывая стабильные трофические состояния, цепи, мы все же склонны к тому, что систематизация будет возможна, У каждого микроорганизма будет свой индивидуальный номер паспорта, но в нем будут отмечены крупные объединяющие блоки, например, национальность: русский или «друг степей калмык».
Представляется необходимым еще раз отметить особенность эволюции микроорганизмов, базирующуюся на уже описанном ранее феномене о возможных длительных периодах стагнации. При изменении экологических условий преимущество получат те или иные микроорганизмы. Однако это не требует выведения из всего пула генетической информации микроорганизмов, не имеющих каких-либо плюсов в данной ситуации. Они остаются и ожидают других экологических перемен, в которых они могут использовать уже накопленную информацию. Для макроорганизмов невозможность продолжения рода, отсутствие потомства неизбежно приводит к смерти особи и элиминации генетической информации (если в дело не вмешаются микробы).
Таким образом, мы должны констатировать, что законы эволюции микроорганизмов отличаются от дарвиновских, имеют свои характерные особенности, вытекающие из самого устройства микроорганизмов и их подчиненности среде существования.
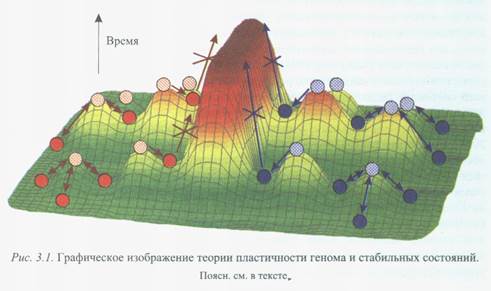
Наше представление эволюции микроорганизмов [Repin et al., 2001], названное теорией пластичности генома и стабильных состояний, можно представить графически (рис. 3.1).
Важной компонентой картинки является вектор времени. Это значит, что все холмы (нестабильные состояния) и долины (стабильные) могут как меняться, так и оставаться в том же виде в зависимости от геофизического состояния планеты или ее частей. Шарики описывают экологические сообщества, среди которых благодаря мутационному процессу, горизонтальному переносу и т. д. могут образовываться варианты, переводящие их в нестабильные состояния на вершины холмов. далее следует переход в то или другое стабильное состояние, существующее на данный момент времени. Естественно, что иногда этот переход требует продолжительного времени, нескольких переходов и не осуществим как однократный акт.
Еще одно важное замечание, которое мы считаем своим долгом сделать: все умозаключения мы даем на том основании, что законы и закономерности, открытые нами, распространяются и на так называемые некультивируемые микроорганизмы, количество которых в настоящий момент на несколько порядков превосходит культивируемую часть. Хотя может быть именно здесь и располагаются микроорганизмы в нестабильных состояниях.
3.2. ТОЧКА ОТСЧЕТА МИКРОБНОЙ ЭВОЛЮЦИИ:
ВОЗНИКНОВЕНИЕ ПЕРВИЧНЫХ ЭКОСИСТЕМ НА ЗЕМЛЕ С ПОЗИЦИИ СИСТЕМНОИ КОНЦЕПЦИИ ЗАРОЖДЕНИЯ БИОСФЕРЫ
Грань между живыми и неживыми природными системами
Неживые органические микросистемы, которые сейчас рассматриваются в качестве предбиологических (макромолекулы мира РНК, липосомы, протеиноидные микросферы и т. д.), и живущие микроорганизмы являются принципиально разными типами природных систем. Любая природная система характеризуется своими собственными свойствами, которые отличают ее от других. Набор ключевых свойств выражает сущность системы. Таким образом, к пониманию грани между живой и неживой природой можно приблизиться через изучение набора свойств, которые свойственны биологическим системам и не свойственны небиологическим, включая предбиологические модели. Этот тезис являлся исходным пунктом для разработки системной концепции зарождения биосферы [Kompanichenco, 2004, 2008; Компаниченко, 2008].
В основу данной концепции положена систематизация и обобщение фундаментальных свойств биологических систем, проведенная через их сопоставление со свойствами систем небиологических. Первоначальный набор был составлен из 230 биологических свойств, выделенных 73 учеными [Fundamentals..., 2002], и является последним обобщением в этой области фундаментальной биологии. Выделенные этими авторами свойства часто повторяют друг друга. Например, способность к самовоспроизведению как одно из главных биологических свойств выделял каждый третий из них. На основе проведенного анализа и обобщения этих данных было сформулировано 31 фундаментальное свойство биологических систем, которые полностью перечислены в [Kompanichenco, 2008; Компаниченко, 2008]. Свойства разделены на две группы. 19 из них отнесены к уникальным фундаментальным свойствам биологических систем, которые не проявляются в существовании любых других природных систем. Согласно авторской интерпретации, они могут быть сведены к четырем интегральным, или ключевым, биологическим свойствам: 1) способность концентрировать свободную энергию и информацию (посредством извлечения из окружающей среды за счет собственной активности); 2) способность оказывать усиленное реагирование на внешние воздействия; 3) целесообразное поведение, или целесообразный характер взаимодействия со средой;
4) регулярное самообновление на различных иерархических уровнях (молекулярном, организменном, видовом, биосферном). Остальные 12 свойств отнесены к неуникальным фундаментальным свойствам биологических систем. Такие же или сходные свойства характеризуют также некоторые неживые системы, хотя в таком случае последние лишены биологической специфичности. Например, кристалл способен накапливать и сохранять информацию о своем росте (одно из неуникальных биологических свойств, выделявшееся несколькими учеными), так же как и живой организм, но он не может ее реорганизовывать и передавать следующим генерациям.
Фактически, уникальные биологические свойства фиксируют отчетливый барьер, разделяющий живую и неживую части природы, в то время как неуникальные могут быть рассмотрены в качестве связующей нити между ними. В течение процесса зарождения жизни на ранней Земле уникальные свойства впервые появились, а неуникальные «перешли» из материнской геологической среды и приобрели биологическую специфику. Следовательно, неуникальные свойства могут служить ориентиром в наших попытках объективно охарактеризовать процессы в геологической колыбели жизни. С этой точки зрения очень примечательными являются такие неуникальные фундаментальные биологические свойства, как термодинамическая и химическая неравновесность, интегрированность, поддерживаемая кооперативными явлениями, и способность к самоорганизации. Необходимость неравновесных условий связывает процесс зарождения жизни с широким кругом специфических явлений, возникающих при неравновесных бифуркационных переходах и формировании диссипативных структур. Они изучаются в рамках теории диссипативных структур и синергетики, основы которых были заложены соответственно И. Пригожиньм и Г. Хакеном вместе с их последователями. Изученные ими универсальные закономерности бифуркационных переходов химических систем были использованы при рассмотрении процессов в геологической колыбели жизни и обосновании перехода органических микросистем (оптимальный состав которых еще предстоит выяснить) в сообщества первоначальных форм жизни ─ пробионтов. Их последующая эволюция привела к формированию более сложно организованных прогенотов (которые можно сопоставлять с last Universal Common Ancector— последним общим предком для всех царств микроорганизмов), и затем архей и бактерий. Таким образом, в соответствии с системной концепцией между доступными сегодня непосредственному изучению предбиологическими моделями и прокариотическими клетками можно теоретически реконструировать несколько промежуточных форм, которые возникали в течение трех этапов зарождения биосферы: пребиотического, переходного и биотического.
|
|
|
