Биотический этап зарождения биосферы
Из излагаемой концепции вытекает, что жизнь могла возникнуть только в не-равновесной жидкой среде, термодинамические и физико-химические параметры которой имеют значительные колебания. Флуктуации параметров среды в оптимальном (вероятно достаточно высокочастотном) режиме необходимы как для поддержания устойчивости бистейт-микросистем (осциллирующих вокруг точки бифуркации), так и в дальнейшем для осуществления способности пробионтов к усиленному и целесообразному ответному реагированию на изменения в окружающей среде. Наиболее удовлетворительно таким требованиям отвечает гидротермальная среда, в которой существуют как относительно стабильные периоды существования, так и мощные макрофлуктуации, которые стимулируются тектономагматичекой активностью планеты. Они сопровождаются колебаниями более низкого ранга. В этом контексте гидротермальные системы и области их разгрузки в океанах и на континентах рассматриваются как наиболее вероятная среда зарождения жизни на Земле. Таким образом, представляется наиболее вероятным появление первичных разрозненных сообществ пробионтов на выходах горячих источников в прибрежных или глубоких областях древнего океана. Сейчас эти ареалы, часто называемые «черными курильщиками», унаследованы сложными гидротермальными сообществами организмов, в основании трофической цепи которых находятся примитивные (гипер)термофильные археи и бактерии. Нельзя исключить также формирования первичных сообществ пробионтов в зонах разгрузки высокотемпературных флюидов в грунтовые воды в областях активного вулканизма на континентах, где также имеют место значительные флуктуации термодинамических и физико-химических параметров. Впоследствии пробионты могли выноситься в океаны с естественным стоком подземных вод (в настоящее время такой сток составляет в среднем несколько кубометров в минуту с одного квадратного километра поверхности). Данные сообщества, которые вначале, вероятно, были изолированы друг от друга, отвечали, по сути, эмбриональной стадии развития первичных локальных экологических систем,
Как промежуточную форму жизни между пробионтами и современными про-кариотами можно рассматривать прогенот, который был теоретически реконструирован С. Woese [19871]. В структуре вымершей части филогенетического древа микроорганизмов он сопоставляется с последним общим предком (Last Universal Common Ancestor) для всех филогенетических ветвей Archaea и Bacteria. Организмы этого типа могли иметь генотип и фенотип (т.е. информация в них сохранялась в фиксированной форме в одном классе молекул и проявлялась в активной форме в другом), но их гены представляли собой физически отдельные единицы, не организованные в линейно упорядоченные структуры. Протеины представляли собой короткие и/или неуникальные последовательности. Это вытекает из того, что протеины нормального размера не могли быть синтезированы без многочисленных ошибок в условиях отсутствия современного уровня точности трансляции. Как следствие, энзимы прогенотов не могли быть такими точными и специфичными, как в современных микроорганизмах. Норма ошибок в механизмах трансляции должна была быть на стадии прогенота на 2—З порядка больше, чем в живущих сегодня клетках. Чтобы поддерживать в этой ситуации минимально функциональные клетки, гены должны были быть разобщены и существовать в большом числе копий. Вероятно, информационной макромолекулой на этой стадии была РНК, функциональная форма нуклеиновых кислот сегодня, не ДНК.
РНК является достаточно сложной макромолекулой, поэтому появление первичного нуклеопротеидного взаимодействия следует отнести к более ранней формы жизни — пробионту. В этот период протекавшие в нем химические процессы уже были вовлечены в организованную сеть метаболических реакций iп vivо, что способствовало резкому повышению термической стабильности нуклеиновых кислот. Так, согласно экспериментальным данным, нуклеиновые кислоты в водной среде in vitro стабильны до температур 50—60 °С (в редких исключительных случаях до 90 °С), в то время как некоторые гипертермофильные археи имеют термостабильные генетические структуры и могут существовать при температуре до 120°С. Изучение соотношений изотопов кислорода в кремнистых известняках океанического происхождения в период древнейших рекордов жизни показало, что температура океана составляла 80 °С 3,8 млрд лет назад и 70°С 3,5 млрд лет назад [Knauth, Lowe, 2003]. Такие же результаты были получены на основе изучения изотопов кремния. Таким образом, температурные условия в гидросфере в период зарождения жизни на Земле (минимум 80°С, а в областях разгрузки гидротермальных систем еще выше), вероятно, не позволяли стабильно существовать даже коротким цепочкам нуклеиновых кислот в неживых пребиотических бистейт-микросистемах, за исключением кратких периодов проблеска жизни при временном положительном F/S-отношении. Большая часть информации в пребиотических бисистемах, вероятно, была сосредоточена в сетях автокаталитических реакций (наличие системы с таким уровнем хранения информации предполагалось и C.Woese). Качественно новые возможности для синтеза полинуклеотидных цепочек появились при трансформации бисистем в пробионты. Термическая устойчивость аминокислот выше, чем нуклеиновых кислот, и в общем случае она возрастает с ростом степени их полимеризации. Таким образом, пробионт, наследуя еще со стадии бистейт-системы колебательный режим существования и поддерживамую им непрерывную циркуляцию веществ и приобретя реверсивную направленность внутренних процессов (F > S), неизбежно должен был вовлекать в процесс собственного строительства различно организованные полимерные цепи аминокислот и
 нуклеотидов, которые функционально дополняли друг друга. В этом процессе должны были участвовать также липиды, которые принимают участие в процессах высокотемпературного геохимического преобразования органических веществ (в частности, при образовании нефти), и ряд других соединений.
нуклеотидов, которые функционально дополняли друг друга. В этом процессе должны были участвовать также липиды, которые принимают участие в процессах высокотемпературного геохимического преобразования органических веществ (в частности, при образовании нефти), и ряд других соединений.
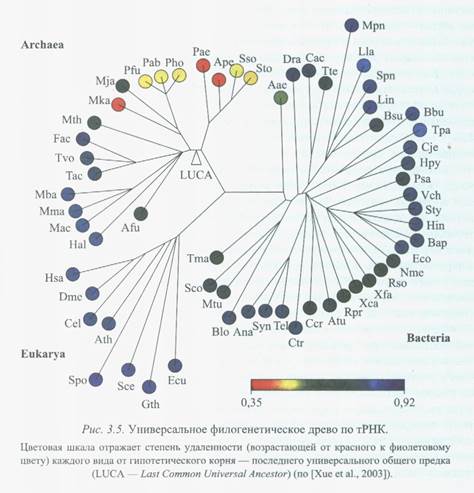
Эволюция прогенотов в прокариотические клетки протекала через значительное увеличение точности трансляции, удлинение цепочек биополимеров обоих классов, появление ДНК и формирование генома, что сопровождалось качественным увеличением структурно-функциональной сложности системы [Woese, 1987]. В экосистемном аспекте этот процесс характеризовался началом видообразования и появлением смежных филогенетических ветвей биологической эволюции. Наличие многочисленных пересекающихся ветвей в эволюционном древе прокариот подчеркивалось, в частности, W.Doolittle [2000]. Согласно наиболее распространенному мнению среди специалистов в микробной экологии, в основании филогенетического древа находятся гипертермофильные или термофильные Archaea и Bacteria [Woese, 1987; Stetter, 1995; Xue et al., 2003], хотя не исключена возможность вторичного возникновения некоторых из этих форм из мезофилов на более поздних этапах биологической эволюции. Это совпадает с приведенными выше оценками температурных условий в гидросфере ранней Земли. Одна из последних версий филогенетического древа микроорганизмов приведена на рис. 3.5. Возникновение видов привело к преобразованию первичных сообществ пробионтов и прогенотов («эмбрионов» экосистем) в иерархически организованные экосистемы микроорганизмов. Их пространственная экспансия привела к охвату всей приповерхностной оболочки Земли и, таким образом, к формированию первичной биосферы планеты.
Воспользуйтесь поиском по сайту:



