 |


Строение и классификация синапсов
|
|
|
|
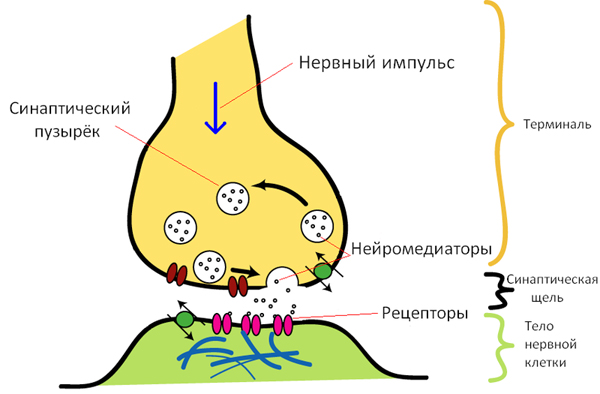
Синапс – это морфофункциональное образование ЦНС, которое обеспечивает передачу сигнала с нейрона на другой нейрон или на эффекторную клетку (мышечную, секреторную). Синапс включает три компонента: пресинаптическая мембрана, постсинаптическая мембрана и синаптическая щель, т. е. содержит элементы и первого и второго контактирующих нейронов.
По локализации синапсы делятся на центральные и периферические. Центральные подразделяются на аксо-аксональные, аксо-дендрические, аксо-соматические, дендро-дендрические и т. д.; по развитию в онтогенезе различают стабильные и динамические синапсы, по конечному эффекту – тормозные и возбуждающие. По механизму передачи сигнала синапсы бывают электрические, химические и смешанные.
Строение синапса
Электрический синапс представляет собой щелевидное образование с ионными мостиками-каналами между двумя контактирующими клетками. При наличии ПД ток почти беспрепятственно перескакивает через щелевидный контакт и индуцирует генерацию ПД в другой клетке, таким образом происходит быстрая передача возбуждения. Но электрические синапсы в основном обладают двусторонней проводимостью. Кроме того с их помощью нельзя заставить эффекторную клетку тормозить свою активность. С другой стороны, передача сигнала происходит почти без синаптической задержки и почти без утечки токов через внеклеточную среду. Электрические синапсы широко распространены в нервной системе беспозвоночных и низших позвоночных. В стволе мозга млекопитающих они имеются в ядрах тройничного нерва и некоторых других ядрах ствола.
Химические синапсы передачу нервного сигнала осуществляют с помощью химических веществ – медиаторов, которые содержатся в синаптических визикулах. Химические синапсы классифицируют по природе медиатора, который они используют: холинэргические (ацетилхолин), адренергические (адреналин), дофаминергические (дофамин) и др.
|
|
|
Классификация синапсов
1. По расположению: центральные: аксосоматические; аксоаксональные; аксодендритные; дендросоматические; дендроаксональные; дендродендритные; соматосоматические; периферические: мионевральные; нейроэпителиальные; синапсы вегетативных ганглиев.
2. Физиологическая классификация - в основе процесс, возникающий на иннервируемой клетке: возбуждающие (деполяризующие) - на иннервируемом органе возникает возбуждение в виде возбуждающего постсинаптического потенциала; тормозные (гиперполяризующие) - на клетке возникает тормозной постсинаптический потенциал.
3. По способу передачи возбуждения через синапс: электрические - с помощью электрического тока, расстояние между нервными волокнами и клетками очень мало; химические - с помощью химических веществ, расстояние между волокном и клеткой - больше. Химические вещества - трансмиттеры (медиаторы). Таких синапсов - большинство.
4. В зависимости от медиатора химические синапсы подразделяются на: холинэргические; адренэргические; гистаминэргические; ГАМК-эргические.
Свойства нервных центров
Нервные центры имеют ряд общих свойств, что во многом определяется структурой и функцией синаптических образований.
1. Одностороннее проведение возбуждения. В ЦНС – в ее нервных центрах, внутри рефлекторной дуги и нейронных цепей возбуждение, как правило, идет в одном направлении – от пресинаптической мембраны к постсинаптической, т. е. вдоль рефлекторной дуги от афферентного нейрона к эфферентному. Это свойство связано со свойствами синапсов.
2. Замедление проведения возбуждения в нервных центрах, или центральная задержка. Замедление проведения возбуждения по нервным центрам получило название центральной задержки. Она обусловлена медленным проведением нервных импульсов через синапсы, так как затрачивается время на следующие выделения медиатора из пресинаптических везикул, выброс его в синаптическую щель и генерация возбуждающего постсинаптического потенциала (ВПСП).
|
|
|
3. Суммация возбуждения и суммация торможения. Принято выделять два вида суммации – временную и пространственную. Временная, или последовательная, суммация проявляется в том, что в области постсинаптической мембраны происходит суммация следов возбуждения во времени, т. е. на нейроне в области его аксонного холмика происходит интеграция событий, разыгрывающихся на отдельных участках мембраны нейрона на определенном отрезке времени. Пространственная суммация возбуждения проявляется в суммировании на аксонном холмике нейрона постсинаптических потенциалов, которые возникают одновременно в различных точках этого нейрона в ответ на приходящие от других нейронов потенциалы действия. Даже если каждый из нейронов в отдельности вызывает лишь подпороговые ВПСП, при синхроннном их появлении они будут способны довести мембранный потенциал в области аксонного холмика нейрона до критического уровня деполяризации и тем самым вызывать возбуждение нейрона. Все сказанное в полной мере относится и к явлению суммации торможения.
4. Явление окклюзии (или закупорки) отражает эффект взаимодействия между собой двух импульсных потоков, при котором имеет место взаимное угнетение рефлекторных реакций. Суммарная ответная реакция (рефлекс), вызываемая одновременным воздействием двух потоков, меньше, чем сумма двух реакций, возникающих при действии каждого из этих двух потоков в отдельности. Согласно Ч. Шеррингтону, явление окклюзии объясняется перекрытием синаптических полей, образуемых афферентными звеньями двух взаимодействующих рефлексов.
5. Явление облегчения, которое по своему внешнему проявлению противоположно окклюзии. 0но проявляется в том, что при совместном раздражениим рецептивных полей двух рефлексов наблюдается усиление реакций организма на действие двух раздражителей одновременно.
|
|
|
6. Трансформация ритма возбуждения. Это одно из свойств нейрона как компонента нейронной цепи, которое обнаруживается в процессе проведения возбуждения по нейронным цепям. Трансформация ритма возбуждения заключается в способности нейрона изменять ритм приходящих импульсов. Она проявляется и в противоположном феномене – частота приходящих к нейрону импульсов выше, чем частота генерации ПД при ответе нейрона на эти импульсы.
7. Последействие. Это одно из свойств, характерных для нейронных цепей. Оно заключается в том, что реакция нейрона (в виде генерации одиночных ПД или пачек ПД) на приходящий к нему импульс продолжается длительное время. В основе этого удивительного эффекта, как предполагают, лежат два механизма. Первый связан с наличием длительного ВПСП, возникающего в ответ на приходящий к нейрону импульс (подобная ситуация характерна и для явления трансформации ритма возбуждения). Нередко это явление называется фасилитацией (облегчение). Второй механизм связан с наличием в ЦНС своеобразных "ловушек возбуждения", по которым происходит длительная (в течение нескольких минут или нескольких часов) циркуляция потока импульсов, называемая реверберацией нервных импульсов.
8. Высокая утомляемость нервных центров. Это свойство характерно для нейронных цепей, в том числе для рефлекторных дуг. С одной стороны, оно проявляется в том, что в нейронных цепях, как и в других многозвеньевых системах, может развиваться утомление, которое проявляется в постепенном снижении (вплоть до полного прекращения) рефлекторного ответа при продолжительном раздражении афферентных нейронов.
9. Тонус нервных центров. Для многих нейронных объединений, или нервных центров, характерна фоновая активность, т. е. генерация нервных импульсов с определенной частотой на протяжении длительного времени. Такая активность обусловлена не наличием в составе данного объединения нейрона- пейсмкера (фоновоактивного нейрона), а постоянным возбуждением афферентного нейрона благодаря непрерывному раздражению сенсорных рецепторов. Тонус нервных центров обеспечивает постоянную импульсацию к соответствующим периферическим системам, а также постоянное межцентральное взаимодействие.
|
|
|
10. Пластичность нервных центров – это их способность к перестройке функциональных свойств и, в определенной степени, функций под влиянием длительных внешних воздействий или при очаговых повреждениях мозга. Посттравматическая пластичность нейронных объединений выполняет компенсаторную (восстановительную) функцию, а пластичность, вызванная длительным афферентным раздражением, – приспособительную функцию. Например, для процесса обучения пластичность нейронных объединений является необходимым условием, т. е. его рабочим механизмом. В целом благодаря свойству пластичности нервный центр может существенно модифицировать течение рефлекторных реакций. Основным фундаментом, позволяющим реализовать свойство пластичности, очевидно, следует считать наличие у каждого нейрона в отдельности огромного числа синаптических связей, а также возможность изменения синтетических процессов внутри каждого нейрона.
Строение спинного мозга
Спинной мозг лежит в позвоночном канале и представляет собой тяж длиной 41-45 см (у взрослого человека среднего роста). Он начинается на уровне нижнего края большого затылочного отверстия, где выше расположен головной мозг. Нижняя часть спинного мозга сужается в виде конуса спинного мозга. Вначале, на втором месяце внутриутробной жизни, спинной мозг занимает весь позвоночный канал, а затем вследствие более быстрого роста позвоночника отстает в росте и перемещается вверх.
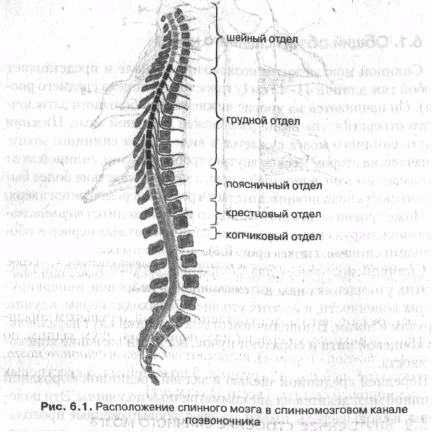
Ниже уровня окончания спинного мозга находится терминальная нить, окруженная корешками спинномозговых нервов и оболочками спинного мозга (рис. 6.1).
Спинной мозг имеет два утолщения: шейное и поясничное. В этих утолщениях находятся скопления нейронов, иннервирующих конечности, и из этих утолщений выходят нервы, идущие к рукам и ногам. В поясничном отделе корешки идут параллельно концевой нити и образуют пучок, носящий название конского хвоста.
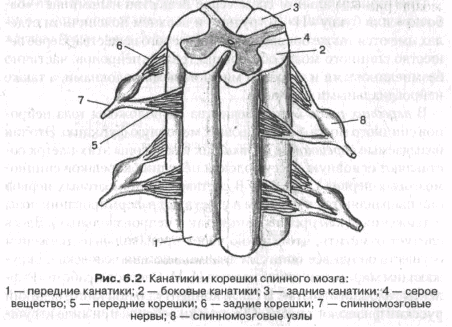
Передней срединной щелью и задней срединной бороздкой спинной мозг делится на две симметричные половины. Эти половины, в свою очередь, имеют по две слабовыраженные продольные борозды, из которых выходят передние и задние корешки, формирующие затем спинномозговые нервы. Благодаря наличию борозд каждая из половин спинного мозга разделена на три продольных тяжа, называемых канатиками: передний, боковой и задний. Между передней срединной щелью и переднебоковой бороздой (местом выхода передних корешков спинного мозга) с каждой стороны находится передний канатик. Между переднебоковой и заднебоковой бороздами (вход задних корешков) на поверхности правой и левой сторон спинного мозга формируется боковой канатик. Позади заднебоковой борозды, по бокам от задней срединной борозды, находится задний канатик спинного мозга (рис. 6.2).
|
|
|
Передний корешок образован аксонами двигательных (моторных) нейронов. По нему нервные импульсы направляются от спинного мозга к органам. Именно поэтому он «выходит». Задний корешок, чувствительный, образован совокупностью аксонов псевдоуниполярных нейронов, чьи тела образуют спинномозговой узел, располагающийся в позвоночном канале за пределами ЦНС. По этому корешку в спинной мозг поступает информация от внутренних органов. Поэтому этот корешок «входит». На протяжении спинного мозга с каждой стороны имеется 31 пара корешков, образующих 31 пару спинномозговых нервов.
Участок спинного мозга, соответствующий двум парам корешков спинномозговых нервов (двум передним и двум задним, по одному с каждой стороны), называют сегментом спинного мозга. Различают 8 шейных, 12 грудных, 5 поясничных, 5 крестцовых и 1 копчиковый сегмент (всего 31 сегмент).
|
|
|
