 |


Свойства потенциала действия
|
|
|
|
Благодаря теории ионного возбуждения Ходжкина — Хаксли, что является существенным доказательством ее правильности, стало возможным объяснить с помощью основных ее положений свойства потенциала действия, а также механизмы его распространения в нервных и мышечных клетках.
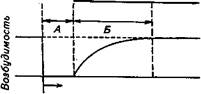
Аккомодация. Закон раздражения Дюбуа-Реймона. Как было выяснено, для того чтобы возник потенциал действия, необходимо деполяризовать мембрану до порогового значения. Однако ни пороговый раздражающий ток, ни пороговый потенциал не имеют определенной величины, поскольку зависят от нескольких факторов, в том числе и от скорости деполяризации мембраны. Зависимость порога возникновения потенциала действия от скорости нарастания раздражающего тока получила название закона раздражения Дюбу а-Р е й м о н а, суть которого состоит в том, что для достижения порога раздражения сила тока должна нарастать с определенной скоростью. При медленном нарастании силы тока возбуждение не наступает (рис. 2.13). Это явление, названное аккомодацией, связано со свойствами инактивации (закрывания) натриевых каналов и активации калиевых каналов при длительной деполяризации. В частности, повышение мембранного потенциала к пороговому уровню вызывает открывание натриевых каналов и соответственно возникновение входящего, натриевого тока. Однако, если скорость этого процесса недостаточна, часть натриевых каналов успевает закрыться. В то же время при деполяризации происходит активация калиевых кана-

|
| ■* Рис. 2.13. Схема, поясняющая аккомодацию в электровозбудимой мембране: А — кривая повышения порога возбуждения; 1...4— нарастание силы раздражающего им- 4 пульса с различной скоростью |
| Время |
лов, у которых в отличие от натриевых инактивация не происходит. Таким образом, ослабляется входящий натриевый ток и усиливается выходящий калиевый ток. Все это замедляет или блокирует развитие регенеративного процесса.
|
|
|
| °0 3,2 |

|
Закон длительности раздражения. Закон Дюбуа-Реймона тесно связан с другим необходимым условием наступления возбуждения — определенной длительности раздражения. Исследованию данной проблемы были посвящены многочисленные работы, начатые еще в конце XIX в., экспериментальные данные которых позволили сделать вывод, что порог возбуждения не может быть достигнут, если раздражение длится очень долго, но сила его мала, или сила раздражения велика, но действует очень короткий промежуток времени. Кривая, выражающая зависимость необходимой силы тока от времени раздражения нервного волокна, имеет вид гиперболы и никогда полностью не приближается к осям координат (рис. 2.14). Начиная с некоторого момента кривая идет параллельно оси абсцисс — оси времени, т. е. сила ниже порого-
О 0,2 0,4 0,6 0,8 1,0 1,2 1,4 1,6 1,8 2,0 2,2 2,4 2,6 Длительность, мс
Рис. 2.14. Кривая «сила — длительность» для возникновения потенциала в нерве:
/: по оси ординат — реобаза, по оси абсцисс — полезное время; //: по оси ординат — двойная реобаза, по оси абсцисс — хронаксия
вой не вызывает возбуждения ни при какой длительности раздражения — время бесполезно. И наоборот, начиная с пороговой силы, время становится значимым (полезным) для раздражения. По существу, «полезное время» — это критическая длительность действия наименьшего тока, способного вызвать возбуждение. Применимо к нервной или мышечной клетке это возникновение потенциала действия. Для того чтобы импульс тока мог изменить мембранный потенциал и вызвать потенциал действия, через мембрану должно пройти некоторое минимальное количество электричества Q, которое, как известно, измеряется как произведение силы тока /на время t: Q = It. Из соотношения следует, что по мере сокращения длительности тока необходимо увеличить его силу, чтобы он сохранял эффективность в качестве раздражителя.
|
|
|
Для импульсов большой длительности существует некоторая минимальная сила тока, достаточная для возбуждения. Более слабый ток из-за того что мембрана в покое имеет определенную проводимость для ионов калия, не способен вызвать необходимое смещение трансмембранной разности потенциалов и запустить регенеративный процесс генерации потенциала действия. Таким образом, «полезное время» может свидетельствовать об уровне возбудимости ткани: чем короче «полезное время», тем более возбудимыми являются ткани и быстрее реагируют на раздражитель. Это положение с начала XX в. успешно используется в клинической и лабораторной практике. Вместе с тем следует, что точное измерение «полезного времени» затруднительно, поскольку в этой части кривой значительные изменения времени соответствуют очень малым изменениям порога раздражения (см. рис. 2.14). Поэтому французским исследователем Лапиком было предложено измерять в качестве порога времени минимальную длительность раздражителя при его силе, равной двум порогам, и при условии, что сила порогового раздражителя измерена при длительностях, превышающих «полезное время». Временные интервалы и значения силы раздражителя получили специальные названия и используются по настоящее время. Пороговая сила была обозначена как реобаза, а необходимое время раздражения при силе раздражения, равной двум реобазам, — хронаксия. Хронаксия — величина переменная и зависит от многих факторов: структуры ткани, ее функционального состояния и всего организма в целом. Кривые «сила — длительность» для разных тканей и органов подобны по форме, но отличаются во времени. Например, у лошади и жвачных животных хронаксия двигательных нервов колеблется от 0,09 до 0,2 мс, а скелетных мышц — от 0,2 до 0,4 мс; самая большая хронаксия у гладких мышц желудка, кишечника и матки, измеряемая десятками и сотнями миллисекунд.
Рефрактерность. После окончания возбуждения в нервных или мышечных клетках или, другими словами, после окончания в них потенциала действия наступает состояние временной невозбуди-
|
|
|

|
Рис. 2.15. Периоды рефрактерности (снижения возбудимости) при генерации потенциала действия:
верхняя запись — потенциалы действия, генерируемые на электрическое раздражение, нижняя запись — отметка электрического раздражения; А — период абсолютной рефрактерности; Б— период относительной рефрактерности; 1...4— периоды рефрактерности
| 'jj[ |
| . Время после первого стимула |
 мости — рефрактернос-т и. Наличие фазы рефрактерности, сопровождающее окончание возбуждения, было впервые установлено в конце XIX в. Мареем на сердце лягушки. Он обнаружил, что после сокращения сердца очередное сокращение нельзя вызвать в течение периода, равного десятым долям секунды, независимо от амплитуды и длительности раздражающего стимула. В нервных клетках период невозбудимости оказался значительно короче, чем в мышечных. При уменьшении интервала раздражения между двумя раздражающими электрическими стимулами величина потенциала действия в ответ на второй стимул становится все ниже и ниже. А если повторный стимул наносится во время генерации потенциала действия или сразу же после его окончания, то второй потенциал действия не генерируется (рис. 2.15). Период, в течение которого потенциал действия на второй раздражающий стимул не возникает, получил название абсолютного рефрактерного периода. Он составляет для нервных клеток позвоночных животных 1,5...2 мс. После периода абсолютной рефрактерности наступает относительный рефрактерный период, который характеризуется: 1) повышенным порогом раздражения по сравнению с исходным состоянием (т. е. для того чтобы возник повторный потенциал действия, необходим ток большей величины); 2) снижением амплитуды потенциала действия. По мере окончания периода относительной рефрактерности возбудимость повышается до исходного уровня и величина порогового раздражения уменьшается также до первоначального значения.
мости — рефрактернос-т и. Наличие фазы рефрактерности, сопровождающее окончание возбуждения, было впервые установлено в конце XIX в. Мареем на сердце лягушки. Он обнаружил, что после сокращения сердца очередное сокращение нельзя вызвать в течение периода, равного десятым долям секунды, независимо от амплитуды и длительности раздражающего стимула. В нервных клетках период невозбудимости оказался значительно короче, чем в мышечных. При уменьшении интервала раздражения между двумя раздражающими электрическими стимулами величина потенциала действия в ответ на второй стимул становится все ниже и ниже. А если повторный стимул наносится во время генерации потенциала действия или сразу же после его окончания, то второй потенциал действия не генерируется (рис. 2.15). Период, в течение которого потенциал действия на второй раздражающий стимул не возникает, получил название абсолютного рефрактерного периода. Он составляет для нервных клеток позвоночных животных 1,5...2 мс. После периода абсолютной рефрактерности наступает относительный рефрактерный период, который характеризуется: 1) повышенным порогом раздражения по сравнению с исходным состоянием (т. е. для того чтобы возник повторный потенциал действия, необходим ток большей величины); 2) снижением амплитуды потенциала действия. По мере окончания периода относительной рефрактерности возбудимость повышается до исходного уровня и величина порогового раздражения уменьшается также до первоначального значения.
|
|
|
Наличие периодов абсолютной и относительной рефрактерности легко объяснимо на основании данных о динамике активации и инактивации натриевых и калиевых ионных каналов во время
генерации потенциала действия. Так, в период абсолютной рефрактерности во время нисходящей части потенциала действия повышается калиевая проводимость за счет открывания дополнительных калиевых каналов и снижается натриевая проводимость за счет инактивации натриевых каналов. Поэтому даже при больших значениях деполяризующего тока не удается активировать такое число натриевых каналов, чтобы выходящий натриевый ток мог бы превысить увеличенный выходящий калиевый ток и снова запустить регенеративный процесс. Во время относительного рефрактерного периода деполяризующий сигнал достаточно большой амплитуды может активировать воротный механизм натриевых каналов так, что, несмотря на большое число открытых калиевых каналов, натриевая проводимость увеличивается и вновь возникает потенциал действия. Вместе с тем из-за увеличенной проводимости мембраны к ионам калия и остаточной натриевой инактивации повышение мембранного потенциала не будет уже столь близко к значению равновесного натриевого потенциала, поэтому потенциал действия будет меньшим по амплитуде.
|
|
|
